In mammals, neural stem cells appear early in development and remain active within the central nervous system for the whole life duration of the organism. During this developmental process they assume different cellular morphologies and reside within changing microenvironments whilst retaining the basic properties of a stem cell: multipotentiality and the ability to self renew. In this chapter, the basic morphological characteristics of neural stem cells will be reviewed, along with the fundamental structural components and signalling molecules of their microenvironments. In early neural development, when the patterning of the nervous system is established, neural stem cells are called neuroepithelial cells; they are situated among other neuroepithelial cells and they are exposed to various signals such as retinoic acid, sonic hedgehog and fibroblast growth factors. When neurogenesis commences, stem cells are transformed to radial glial cells and the complexity of their microenvironment increases due to the emergence of various types of neuronal progenitors, differentiated cells and extracellular signaling molecules. Finally, during adulthood, neural stem cells assume astroglial morphology and reside in specific microenvironments that are called neurogenic niches; small neurogenic islands where neurons and glia are continuously generated under the control of mechanisms largely similar to those operating during embryonic development.
1. The embryonic neural stem cell (NSC) microenvironment
Central nervous system (CNS) development is an intricate process relying on a series of mechanisms precisely regulated in time and space. In rodents, the majority of the cells present in the adult brain are produced and migrate to their respective destination within an approximately one-week period during embryogenesis. The embryonic CNS is a dynamic structure, constantly increasing in size due to histogenesis, while the stem/precursor cell populations which are responsible for building the brain are retained in two distinct and relatively small proliferative areas. The first is the ventricular zone (VZ) where epithelial cells with NSC properties appear approximately at embryonic day (E) 8 and from which originate all cells of the developing and mature CNS, including adult NSCs (Alvarez-Buylla etal., (2001)). After a period of NSC/precursor expansion, as neurogenesis commences, a second progenitor population starts to be generated from asymmetrically dividing cells in the VZ and migrates basally. These cells, termed intermediate progenitors or basal progenitors, divide symmetrically to produce neurons and glia. They are found throughout the CNS and in the telencephalon the region containing these cells is called subventricular zone (SVZ; Martinez-Cerdeno etal., (2006); Smart, (1972), 1973).
1.1. The VZ microenvironment
The early NSC microenvironment seems to be homogeneous in terms of cellular morphologies (Pinto and Gotz, (2007)). It consists of characteristic bipolar cells, termed neuroepithelial (NEP) cells, with one process (apical) attached to the ventricle and one longer process (basal) attached to the pial surface. NEP cells form the pseudostratified VZ and are characterized by the periodic apico-basal translocation of their nucleus (interkinetic nuclear migration) which is regulated in such a way that mitosis occurs always at the ventricular surface, while S phase occurs at the basal-most area (Gotz and Huttner, (2005); Pinto and Gotz, (2007)). The main structural constant of this early microenvironment is the ventricle, while the thickness of the neuroepithelium increases with time, accommodating the augmenting number of NEP cells and the occasionally generated neurons that quickly migrate towards the pial surface of the nervous tissue. Around midgestation, in rodents, NEP cells start to express glial markers and assume a more elongated morphology. Reflecting this transition, the emerging neural stem cell/progenitor type is now named a radial glial cell (RG) and retains the bipolar morphology and the interkinetic nuclear migration characteristic of NEP cells. As the thickness of the nervous tissue increases with the generation of large numbers of neurons, the basal process of the RG elongates in order to retain attachment to the pial surface (Rakic, (2003)). Therefore, RG appear to be the only embryonic CNS cells able to sense and integrate information from at least four different microenvironments: i) the VZ, consisting mainly of RG cell bodies and apical processes as well as newly generated neurons migrating away; ii) the SVZ, consisting of basal progenitors and newly generated neurons/glia; iii) the mantle, consisting of post-mitotic cells; and iv) the basal lamina of the pial surface, an extracellular matrix (ECM) rich membrane contacted by RG basal process. Although it remains unclear whether extracellular signalling communication exists among these microenvironments, a recent study revealed the presence of a neuron-secreted cytokine (cardiotrophin 1; Barnabe-Heider etal., (2005)) responsible for the initiation of gliogenesis.
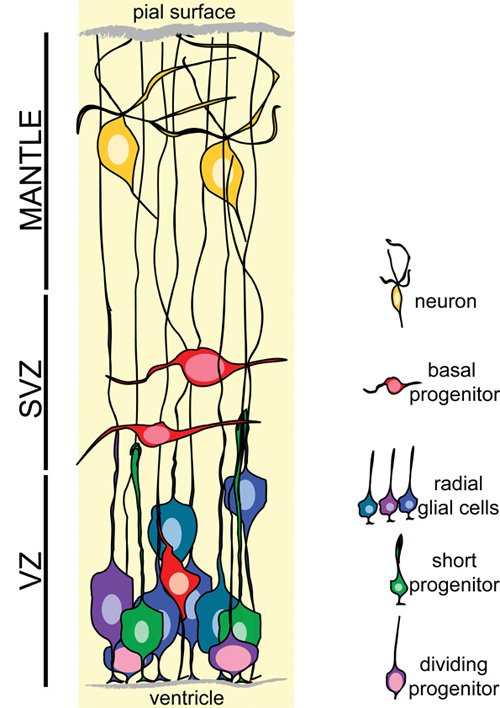
Figure 1. The embryonic neural stem cell microenvironment.
The complexity of the mantle microenvironment increases as development proceeds, due to the generation of different neuronal cell types and the appearance of a dense network of blood vessels (Herken et al., (1989)). A parallel increase in the VZ complexity is observed (Pinto and Gotz, (2007)), although this is not structurally obvious because the VZ is dominated by bipolar, “identical” RG with only a few intercalated short progenitors (Gal et al., (2006); Hartfuss et al., (2003)). However, immunostaining with different markers combined with results from fate studies suggest the coexistence of RG sub-types with different functions and lineage commitment (Hartfuss et al., (2003); Hartfuss et al., (2001); Malatesta et al., (2000); Plachta et al., (2004); Williams and Price, (1995)) and it is well established that RG carry different intrinsic information depending on their positional characteristics (dorsal-ventral, rostral-caudal; Guillemot, (2005)). Finally, to add to the complexity of the progenitor microenvironments, it should be noted that specific areas secreting growth factors or morphogens exist (named signalling centers) either within the VZ/SVZ or outside (Assimacopoulos et al., (2003); Shimogori et al., (2004)).
1.2. Signalling in the embryonic NSC microenvironment
1.2.1. Intrinsic regulation
The behavior of NSC/precursor cells is controlled by both extrinsic and intrinsic mechanisms. Clonally grown E10 cortical progenitors generated first neurons and then glia, similar to the way neurogenesis precedes gliogenesis in vivo (Qian et al., (1998); Qian et al., (2000)) and a follow-up study revealed that this intrinsic timer extended to the “correct” sequential generation of different cortical neuron subtypes (Shen et al., (2006)). In addition, transplantation experiments have revealed that progenitors maintain their intrinsic potential when grafted ectopically (Darsalia et al., (2007); Olsson et al., (1998)). Many transcription factors have been shown to play a role in NSC/progenitor cell proliferation and/or differntiation. These include proneural genes encoding for basic-helix-loop-helix (bHLH) transcription factors (Bertrand et al., (2002)), SRY-related HMG box (SOX) family transcription factors (Episkopou, (2005)), the nuclear receptors estrogen receptor (Brannvall et al., (2002)), peroxisome proliferator activated receptor γ (Wada et al., (2006)) and N-CoR, a nuclear receptor co-repressor (Hermanson et al., (2002)). Loss or gain of function of many of the above factors has been shown to be sufficient to alter progenitor specification, cell cycle and fate irrespective of the environment (Bertrand et al., (2002); Campbell, (2003); Guillemot, (2005)). Another mechanism implicated in the control of cell intrinsic properties is epigenetic modifications. Epigenetic regulation involves histone and DNA modifications that alter the condensation of the chromatin and thus the activity of genes. Several types of histone modification have been described including methylation, acetylation and phosphorylation. Histone acetylation of neuronal gene promoters is modulated by histone acetyltransferases (HATs) and deacetylases (HDACs) and is essential for the repression of these genes in undifferentiated neuronal progenitors (Ballas and Mandel, (2005)). Furthermore, proliferating NSCs have different patterns of histone methylation compared to differentiated neurons (Biron et al., (2004)). Finally, it is believed that microRNAs also act as intrinsic regulators of neuronal progenitor behavior (Cao et al., (2006)), a field where rapid progress can be expected in the near future.
1.2.2. Diffusible signals
The microenvironment (extrinsic factors) can regulate the behavior of neuronal progenitors through diffusible signals and/or molecules mediating cell to cell and cell to ECM interactions. Before examining individual diffusible signalling molecules, it should be emphasized that these molecules, by their nature, form gradients within the tissue and can signal in areas distant from their sources. Therefore, in each position within the developing CNS, a neural stem cell/ progenitor would be exposed to a unique combination of signals that might instruct appropriate region-specific behavior.
A major group of molecules with key roles in the regulation of cell proliferation and differentiation is growth factors. Several bone morphogenetic proteins (BMPs), members of the transforming growth factor-β (TGFβ) family, are expressed along the dorsal midline of the developing brain and are necessary for midline development (Bertrand and Dahmane, (2006); Campbell, (2003)). Overexpression of BMP 2 and 4 results in decreased cell proliferation and premature neuronal differentiation (Li et al., (1998)) and this effect can be reversed by the addition of the BMP signalling inhibitor noggin (Li and LoTurco, (2000)). In vivo overexpression of truncated BMP type I receptor has provided additional evidence for the role of BMPs in promoting cell differentiation at the expense of proliferation (Li et al., (1998)).
In addition, several fibroblast growth factors (FGFs), and in particular FGF8 and FGF3, are expressed in the anterior neural ridge, the midbrain-hindbrain barrier and the prospective rhombomere 4 of the hindbrain early in development (Mason, (2007)). FGF expression profile increases in complexity during development, and varies between the mouse, chick and zebrafish. FGF activity is critical not only for the patterning of the nervous system but also for other functions, such as the FGF8-dependent survival of cells in the forebrain (Storm et al., (2003)) and the midbrain-hindbrain region (Chi et al., (2003)). Moreover, FGF-2 (or bFGF) is a mitogen widely used in vitro, as it is essential for keeping precursors of the embryonic telencephalon and neural tube in a progenitor state (Kalyani et al., (1997); Kilpatrick et al., (1993); Murphy et al., (1990); Vescovi et al., (1993)) and bFGF deficient mice have small brains due to a marked decrease in the NSC/precursor population (Vaccarino et al., (1999)). The role of the FGF family in embryonic stem/precursor cell behavior is also supported by the reported expression of all FGF receptors (FGFRs) in vivo (Bansal et al., (2003)). FGFR4 was shown to be highly expressed in rat neural tube neuroepithelial cells (Kalyani et al., (1999)), while rat dorsal telencephalon neuroepithelial cells were found to express mostly FGFR1 and 3 with these receptors being pivotal for self-renewing symmetric cell divisions (Maric et al., (2007)).
In addition to growth factors, another group of diffusible molecules that have been implicated in the regulation of neural precursor behavior are morphogens, with sonic hedgehog (Shh) being the prominent example. Shh signalling is mediated by its receptor patched (PTC1), a transmembrane protein (Dessaud et al., (2007)), which in the absence of Shh constitutively represses the G-protein coupled receptor smoothened (Smo). After binding with Shh, PTC1 relieves its inhibition on Smo thus activating downstream signalling pathways which result in the modulation of the transcriptional activators Gli1-3, and the Gli repressor. Both Gli 2 and 3 are strongly expressed in the VZ of mice, while Shh and Gli 1 are weakly expressed (Dahmane et al., (2001); Hui et al., (1994)). Shh is involved in ventral patterning throughout the developing CNS (Bertrand and Dahmane, (2006); Campbell, (2003)), as well as in regulating progenitor proliferation, since Shh deficient mice are characterized by reduced brain size and cyclopia (Chiang et al., (1996); Muenke and Cohen, (2000)). This phenotype is likely to be due to aberrant cell proliferation in both the VZ and SVZ, based on observations in mice deficient for Gli 2 (Palma and Ruiz i Altaba, (2004)). A second morphogen known to be important during CNS development is retinoic acid (RA). It is a diffusible molecule that is produced intracellularly by retinaldehyde dehydrogenases (RALDH1-3). RA is sequestered in the cytoplasm by cellular-RA-binding proteins (CRABP1-2) and acts in the nucleus after binding to RA receptors (RAR1-3) and retinoic X receptors (RXR1-3; Maden, (2002)). RA is important in the very early neuronal microenvironment, where it participates in the regulation of the anterio-posterior axis (Maden, (2002)). In later stages, RA signalling is important for dorso-ventral patterning of the spinal cord (Pierani et al., (1999)) and for patterning of the hindbrain (Marshall et al., (1992)). More recent work has revealed that RA is present in relatively high levels in the ventral forebrain (Takahashi and Liu, (2006)), regulating the specification of the intermediate area between cortex and striatum.
Finally, the Wnt signalling pathway has also been shown to regulate cell behavior in the developing brain. High-level expression of Wnt receptors Frizzled 5, 8, 9 and of secreted frizzled protein 1 have been reported in the VZ of mouse embryos (Kim et al., (2001); Van Raay et al., (2001)), while functional studies have revealed a critical role for Wnt signalling in dorsal forebrain specification (Gunhaga et al., (2003); Hirabayashi et al., (2004); Machon et al., (2007)) and in the regulation of the NSC/precursor cell cycle in the VZ. When β-catenin signalling (the central mediator of the canonical Wnt pathway) was enhanced in vivo, a reduction in cell cycle exit was observed in the VZ leading to an enlarged brain (Chenn and Walsh, (2002)). Furthermore, deletion of β-catenin in cortical progenitors resulted in decreased proliferation and migration defects (Backman et al., (2005); Machon et al., (2003)), and targeted inhibition of β-catenin forced VZ cells to prematurely exit the cell cycle and to differentiate into neurons (Woodhead et al., (2006)). A recent paper suggested that β-catenin signalling is necessary to maintain the VZ progenitor population and is downregulated when VZ progenitors are transitioning towards an intermediate progenitor (SVZ) fate; sustained β-catenin activity resulted in the expansion of the VZ progenitor pool and inhibited the production of intermediate progenitors (Wrobel et al., (2007)).
1.2.3. Cell to cell interactions
Both the VZ and SVZ are characterized by a high density of cell bodies and processes, and cell to cell interactions are therefore likely to be another avenue for the regulation of progenitor behavior. Within the VZ, expression of Ephrins B1 and A5 and Eph A4 and A7 receptors has been reported (Depaepe et al., (2005); Greferath et al., (2002); Mackarehtschian et al., (1999); Stuckmann et al., (2001)). Ephrin B1, in particular, is thought to promote cell migration out of the VZ as its expression forms an apico-basal gradient (Stuckmann et al., (2001)). Ephrin A5/Eph A7 may control the size of the VZ progenitor pool by promoting apoptosis of NSC/precursors (Depaepe et al., (2005)). Cell-cell signalling via the Notch pathway is also known to play a critical role in CNS development as evidenced by a series of mouse mutation studies (Hitoshi et al., (2002); Yoon and Gaiano, (2005)). Several studies have shown expression of components of the Notch pathway in the mouse VZ, such as Notch-1 (Gaiano et al., (2000)), Notch-3 (Dang et al., (2006)) and Delta-1 (Beckers et al., (2000)). Strong expression of Notch-1 and Delta-1 and weak expression of Notch-3 has been reported in the human VZ (Kostyszyn et al., (2004)). Moreover, intraventricular injection of Notch ligands increases the numbers of newly-generated precursor cells (Androutsellis-Theotokis et al., (2006)). Notch signalling also plays a critical role in cell fate determination since activation of either Notch-1 or Notch-3 results in increased numbers of radial glia (Dang et al., (2006); Gaiano et al., (2000)). Consistent with these data, a decrease in the size of the VZ and premature differentiation has been reported in delta-like-1 deficient mice (Yun et al., (2002)). Finally, a recent study revealed that the response to Notch via the Notch effector CBF-1 is different between the two types of stem/precursor cells of VZ/SVZ, with NSCs but not intermediate progenitors utilizing CBF-1 (Mizutani et al., (2007)).
Another form of cell to cell interaction in the VZ might be mediated by cadherin-dependent adherens junctions (AJs), but little evidence exists for cadherin signalling in the SVZ (Lathia et al., (3267)). The cell layer adjacent to the ventricle is characterised by strong cadherin expression in the apico-lateral part of the cell membrane (Aaku-Saraste et al., (1996)) and recent work has proposed that the behavior of NSC/precursor cell progeny is dependent on the symmetric or asymmetric inheritance of AJs upon division (Kosodo et al., (2004)). There is little functional evidence directly linking cadherin signalling to NSC/precursor behavior in the mouse and the chick because interference with cadherin signalling leads to extensive disruption of the neural tube (Kadowaki et al., (2007); Radice et al., (1997)). However, there is experimental evidence suggesting that N-cadherin inhibition leads to hyperproliferation of neural stem/precursor cells (Lele et al., (2002); Noles and Chenn, (2007)). Gap junctions are an additional cell to cell communication system that might be important in the regulation of stem/precursor cell behavior. Connexins (Cxs) 26 and 43 are expressed in the VZ (Bittman and LoTurco, (1999)) and Cx43 has been linked with the ability of bFGF to maintain stem/precursor cells in an undifferentiated state in vitro, potentially by controlling the diffusion of growth factors between cells (Cheng et al., (2004)). Interestingly, inhibition of Cx43 resulted in premature differentiation and cell death even in the presence of bFGF (Cheng et al., (2004)). Gap junctions have also been implicated in precursor migration as mice deficient in Cx43 show an accumulation of precursors in the intermediate zone due to their inability to migrate into the cortical plate (Fushiki et al., (2003)). A recent follow-up study further resolved this effect by showing an unexpected role for Cx26 and 43 in maintaining adhesion of migrating precursor cells on radial glia fibers (Elias et al., (2007)).
1.2.4. Cell to ECM interactions
The ECM is another component of the VZ/SVZ microenvironment that might be crucial in the regulation of NSC/precursor behavior. The cell bodies and the short apical processes of the neural progenitors are positioned in an area devoid of a classic basement membrane but still rich in matrix molecules such as various laminin chains (Campos et al., (2004); Hunter et al., (1992); Lathia et al., (Lathia, J. D. Patton, B. Eckley, D. M. Magnus, T. Mughal, M. R. Sasaki, T. Caldwell, M. A. Rao, M. S. Mattson, M. P. ffrench-Constant, C. (2007a). Patterns of laminins and integrins in the embryonic ventricular zone of the CNS. J Comp Neurol 505, 630–643. Abstract)), the laminin receptor beta1 integrin (Campos et al., (2004); Graus-Porta et al., (2001); Hall et al., (2006); Nagato et al., (2005)), the glycoprotein tenascin-C (Garcion et al., (2004)) and chondroitin sulfate proteoglycans (CSPGs; von Holst et al., (2006)). In addition, many of the bipolar (neuroepithelial or radial glial) cells of the VZ extend a basal process that makes contact with the ECM-rich basement membrane of the pia. Alterations in this basal process microenvironment have been correlated in humans with cortical malformations caused by migration defects (Bonneau et al., (2002); Toda et al., (1994); Yoshida et al., (2001)). Mice deficient for components of the pial ECM such as laminin γ1, integrin α6 or β1 (the α6β1 heterodimer is the principal laminin receptor in the CNS) and reelin have several common defects, such as ectopic growths in the cortical marginal zone and retraction of radial glia basal endfeet (Beggs et al., (2003); Georges-Labouesse et al., (1998); Hartmann et al., 1998; Niewmierzycka et al., (2005)) but without any perturbations in progenitor proliferation or cell fate determination (Haubst et al., (2006)). Functional studies investigating the role of beta-1 integrin in NSC/precursors using in vitro neurosphere forming assays clearly do not allow studies of normal cell to cell and cell to ECM interactions in NSC, and have not provided clear insight on other roles of beta-1 integrin in these cells; blocking of beta-1 integrin, using antibodies, resulted in compromised NSC maintenance (Campos et al., (2004)), but subsequent experiments using neurospheres grown from cells lacking beta1 integrin revealed no similar defects (Leone et al., (2005)). The selective enzymatic degradation of CSPG glycosaminoglycans using chondroitinase ABC in similar in vitro assays resulted in perturbed proliferation and neuronal differentiation, revealing a role of these ECM molecules in the regulation of NSC/precursor behavior (Sirko et al., (2007)).
1.2.5. Blood vessels and cerebrospinal fluid (CSF)
The VZ/SVZ microenvironment is influenced by both the CSF and the numerous blood vessels that begin to form in early stages of CNS development. Blood vessel formation in mice begins as early as E9 (Herken et al., (1989); Vasudevan et al., (2008)) and evidence suggests that neurogenesis and angiogenesis are regulated by common signals, including vascular endothelial growth factor (VEGF), Notch and Shh (Carmeliet, (2003)). An in vitro study revealed that co-culture of E10 neural stem/precursor cells with endothelial cells resulted in larger clones and fewer neurons due to an increase in symmetric, proliferative divisions (Shen et al., (2004)) but the factors responsible remain unknown. The composition and role of the CSF in development remains relatively unexplored. In a recent study CSF flow was perturbed in vivo in chick embryos, resulting in aberrant cortical development (Mashayekhi and Salehi, (2006)), while other experimental work has shown that CSF-derived factors can regulate neural stem/precursor cell behavior in vitro (Gato et al., (2005); Miyan et al., (2006)). Recent work has also shown shedding of prominin-rich vesicles in the CSF, by dividing neural stem cells and that these may provide additional cues, although any such instructive role remains to be determined (Marzesco et al., (2005)).
2. The adult neural stem cell (NSC) microenvironment
During early post-natal life the majority of the remaining neuronal precursors exits the cell cycle and terminally differentiates, mostly into astrocytes and oligodendrocytes. In the rodent brain, this tissue-maturation process is characterized by the gradual shrinkage of the ventricular and subventricular zones and leads to the formation of two highly specialized areas where neurogenesis persists into adulthood: the subependymal zone (SEZ) of the lateral walls of the lateral ventricles and the subgranular zone (SGZ) of the dentate gyrus in the hippocampal formation. Both these areas are embedded in the brain tissue, without any obvious anatomical barrier to separate them from the differentiated cells (in humans the SEZ is bordered by a myelin-rich zone (Curtis et al., (2007))) and continuously contribute new neurons to the olfactory bulb (the SEZ; Doetsch et al., (1999); Goldman and Nottebohm, (1983)) and the granule cell layer of the dentate gyrus (the SGZ; Seri et al., (2001); Seri et al., (2004)).
2.1. The SEZ microenvironment
The SEZ is a thin area beneath the ependymal cell layer that contains the three cell types of the lineage that generates new neurons and glia: infrequently dividing neural stem cells (NSCs), rapidly dividing transit amplifying precursors (TaPs) and neuroblasts (NBs). NBs form clusters all along the ventricular length and migrate in chains towards the dorsal and posterior tip of each lateral ventricle in order to continue their migration within the rostral migratory stream up to the olfactory bulb glomeruli. These three cell types are segregated from the adjacent differentiated brain tissue, forming a cell-dense neurogenic microenvironment; therefore, the SEZ has often been characterized as a stem cell niche (Riquelme et al., (2008)). NSCs have an astrocytic morphology and are situated adjacent to the multiciliated ependymal cells that line the lateral ventricles. They are surrounded by other astroglial cells, TaPs and NBs. In addition, two multicellular structures are integrated into the SEZ: the astrocyte-constructed tubes in which NB clusters migrate towards the rostral migratory stream and the numerous blood vessels with their endothelial cell/pericyte-derived walls and astrocyte endfeet on their surface. Interestingly, the SEZ extracellular matrix (ECM) seems to be significantly different from that of the surrounding mature tissue. Recently it was described that extensions of the vessel basal lamina intrude the SEZ and branch around NSCs and progenitors (Mercier et al., (2002)). These laminin and collagen I-rich ECM structures can be observed under the electron microscope and have been named fractones. Other ECM molecules that have been shown to be present in the SEZ are matrix metalloproteinases, brevican (Jaworski and Fager, (2000)), tenascin-C (de Chevigny et al., (2006); Jaworski and Fager, (2000); Kazanis et al., (2007)), chondroitin/dermatan sulfate proteoglycans (Akita et al., (2008); von Holst et al., (2006)), as well as the trisaccharide LeX/SSEA-1/CD15 that is expressed on NSCs and TaPs and is shed in the microenvironment (Capela and Temple, (2002)). The expression of most of these molecules is normally downregulated during early post-natal life resulting in the formation of the classic brain parenchymal ECM, characterized by the dominance of proteoglycans like brevican, neurocan and versicans (Bandtlow and Zimmermann, (2000); Novak and Kaye, (2000); Rauch, (1997); Ruoslahti, (1996)). Therefore, the distinct nature of the SEZ niche is emphasized by the persistent presence of ECM molecules that are expressed during embryonic development or after injury (Bandtlow and Zimmermann, (2000); Rauch, (1997); Thomas et al., (1996)).
2.2. The SGZ microenvironment
The neurogenic area of the hippocampus is restricted to the thin subgranular zone, near the dentate gyrus hilus. Within this zone, cells divide occasionally in order to produce immature granule neurons, again through an intermediate progenitor stage. While there is some debate about whether these cells are restricted to the production of neurons or are multipotent, and whether their self-renewing capacity is limited (Becq et al., (2005); Bull and Bartlett, (2005); Seaberg and van der Kooy, (2002)), they do continue to divide and produce new neurons throughout life as would be expected of a true stem cell. The newborn daughter cells migrate short distances within the granule cell layers until they reach a final position and start to integrate into the hippocampal formation (Seri et al., (2004)). The SGZ neurogenic system does not form an anatomically separate structure as intermediate progenitors and immature neurons are immersed in the granule cell zone, and little is known about the expression of specific ECM molecules. However, the SGZ retains a specific functional role and thus has still been referred to as a neurogenic niche, with the NSC surrounded by intermediate progenitors, astroglial cells, mature granule cells and resting in close proximity to blood vessels, similarly to the SEZ (Palmer et al., (2000)).
2.3. Signalling in the NSC microenvironment: Structural aspects
The stem cell niche is defined as a microenvironment that facilitates the survival and self renewing capacity of the stem cells, as well as (in the adult CNS) the production of actively dividing precursors leading to the generation of post-mitotic progeny. Contact of the stem cell with adjacent supporting cells is thought to be important, as exemplified by the instructive role of the hub cells in the drosophila germ cell niches (Chen and McKearin, (2005); Fuller and Spradling, (2007)). These germ cells retain their function as long as they remain anchored to the hub cells and their divisions occur in such a way that one daughter cell keeps its contact with the hub cell while the other one loses this contact and proceeds to generate oocytes or spermatocytes. Similar instructive relations have been described in adult mammalian stem cell niches, for example between hematopoietic stem cells and neighboring osteoblasts (Yin and Li, (2006)), or epithelial cells and intestinal stem cells (Walker and Stappenbeck, (2008)).
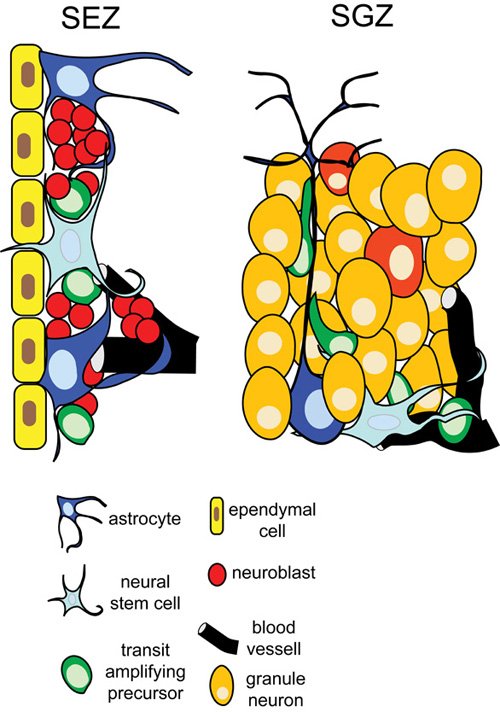
Figure 2. The adult neural stem cell microenvironment in the subependymal zone (SEZ) and the subgranular zone (SGZ).
At present there is no direct evidence to suggest the unique importance of any single interaction between the NSC and the cellular or parenchymal components of the SEZ or SGZ microenvironment. However, the physical positioning of the NSC adjacent to the ependymal cells implies an interaction similar to the hub cell-SC interaction in Drosophila. Ependymal cells exert a supporting/ regulatory function in the niche, since they can modulate the transport of ions and other factors from the cerebrospinal fluid (CSF; Bruni, (1998)). They are also a local source of neurogenic factors like pigment epithelium-derived factor (PEDF; Ramirez-Castillejo et al., (2006)) and the pro-neurogenic BMP signalling modulator noggin (Lim et al., (2000); Peretto et al., (2004)), and they form gap junctions with SEZ astrocytes (Zahs, (1998)). These factors may be required for the maintenance of neural stem cells. Indeed, as discussed earlier, ependymal cells are absent from the SGZ, this being the most distinct structural difference between the two adult neurogenic niches, and there is evidence suggesting that the ependyma-free SGZ contains neuronal progenitors with restricted self-renewing capacity rather than NSCs (Becq et al., (2005); Bull and Bartlett, (2005); Seaberg and van der Kooy, (2002)). In addition, the constant movement of the ependymal cilia is thought to contribute to the generation of gradients of soluble factors in the CSF and to regulate the migration of NBs (Sawamoto et al., (2006)). Therefore any migratory cues provided by ependymal cells would be absent in the SGZ. However, SGZ progenitors do not migrate long distances and can probably acquire the necessary directional cues from the radial processes of the SGZ astrocytes (Seri et al., (2001); Seri et al., (2004)). Nevertheless, a recent study demonstrated that neurogenesis can be maintained in the SEZ after ablation of the ependymal cell layer (Del Carmen Gomez-Roldan et al., (2008)) although the investigation did not address any long-term effects of the ependymal loss.
Three other cell types may also regulate NSC behaviour. In vitro data support the conclusion that the interaction between NSCs and blood vessel endothelial cells might be important in neurogenesis (Shen et al., (2004)). Actively dividing cells have been shown to be positioned near blood vessels in the SGZ (Palmer et al., (2000)) and in the SEZ (Kerever et al., (2007)) and endothelial cells are a source of factors that have been suggested to control neurogenesis, like PEDF, leukemia-inhibitory factor and brain-derived neurotrophic factor (BDNF; Riquelme et al., (2008)). As described earlier, NSCs are also in close contact with their progeny. The induction of massive NSC mitotic activity in the SEZ after ablation of TaPs and NBs by intracerebrally infusing the anti-mitotic drug AraC (Doetsch et al., (1999)), indicates the existence of progenitor-dependent feedback loops controlling NSC proliferation, although the nature of this signalling remains elusive. Astroglia are the most abundant cell type in the SEZ but it is still unknown whether they can be segregated into distinct functional groups, such as astroglia with structural, supporting or neurogenic roles. Astroglia of the SGZ can be structurally separated into radial and horizontal astrocytes (Seri et al., (2004)), while SEZ astroglia (or type-B cells) into two classes, type B1 and B2. B1 astrocytes reside adjacent to the ependymal cells and proliferate less than the smaller and basally located B2 astrocytes (Doetsch et al., (1997)). Using various markers distinct pools of astrocytes have been described in the SVZ, either with NSC properties (expressing PDGFRα- or LeX; Capela and Temple, (2002); Jackson et al., (2006)) or with structural roles (tenascin-C expressing astrocytes at the border of the SEZ or forming the NB migration tubes; Kazanis et al., (2007)). Astrocytes are coupled with gap junctions and are thus able to form a network and transport information from distant areas (Giaume and Venance, (1998)). Thus they act as sensors and modulators of the microenvironment that become reactive, after AraC-induced depletion of the SEZ progenitors even before the mitotic activation of NSCs (Kazanis et al., (2007)). Finally, it is important to note that a potentially important interaction exists between the NSC and the ventricular environment, as NSCs of the SEZ extend a monocliated process in between the ependymal cells enabling them to “taste” the growth factor and morphogen-rich CSF (Alvarez-Buylla et al., (2001); Doetsch et al., (2002)). The importance of cilia in several signalling mechanisms has been recently highlighted (Singla and Reiter, (2006)) and previous experimental work has revealed that essential components of the Shh signalling pathway are positioned at the primary cilium (Corbit et al., (2005); Rohatgi et al., (2007)). In the adult CNS, the significance of primary cilia in neurogenesis was highlighted by the finding that when the cilium was genetically ablated, proliferation in the SGZ was largely compromised (Han et al., (2008)).
2.4. Signalling in the NSC microenvironment: Molecular aspects
2.4.1. Diffusible signals
Cells within the adult neurogenic niches rely on growth factor signalling, cell to cell contact, and cell to ECM interactions for homeostatic cell turnover and increased cell production in response to stimulation (such as injury). Growth factors can originate from cells within the niche or from external sources, mainly the CSF and blood vessels. An increase in SEZ cell proliferation has been reported after infusion of FGF, EGF, and TGFalpha (Craig et al., (1996); Kuhn et al., (1997); Wagner et al., (1999)), all of which could originate from the CSF (especially from the choroid plexus), and have no identified sources within the niche. It is worth noting that in these early studies NSC and progenitor populations were analyzed together, not allowing the identification of specific cell types. In the case of EGF, a subsequent study (Doetsch et al., (2002)) revealed that the EGF-induced proliferation in the SEZ was driven largely by TaPs. A number of vasculature-related growth factors have been demonstrated to regulate NSC and progenitor proliferation, most notably VEGF, PDGF and PEDF. SEZ progenitor proliferation can be enhanced by VEGF both in vitro and after intraventricular administration (Jin et al., (2002)), while this growth factor is believed to mediate the exercise-dependent upregulation of proliferation in the SGZ (Cao et al., (2004)). NSCs also express the PDGFalpha receptor, with the level of expression maintaining the balance between neuron and oligodendrocyte production. Conditional ablation of the receptor in SEZ NSCs did not affect neurogenesis but did result in a reduction in oligodendrocyte production, while intraventricular infusion of PDGFalpha into the lateral ventricles increased type-B cell production and blocked neuroblast generation (Jackson et al., (2006)). PEDF is another factor produced by endothelial cells (and ependymal cells) that was shown to selectively activate NSCs after intraventricular infusions (Ramirez-Castillejo et al., (2006)). Finally, other factors secreted by blood vessels that are known to influence NSC/progenitor behaviour are leukaemia-inhibitory factor (LIF) (Mi et al., (2001)) and brain-derived neurogenic factor (BDNF; Leventhal et al., (1999); Scharfman et al., (2005)). Bone morphogenetic proteins (BMPs) and their receptors are expressed in the SEZ (Colak et al., (2008); Lim et al., (2000)). In agreement with data from embryonic development, they were thought to direct progenitors towards a glial lineage, a function antagonised in the SEZ by the ependyma-derived noggin (Lim et al., (2000)). However, a recent study revealed that the role of BMPs in the adult neurogenic process is more complicated since conditional deletion of the BMP signalling mediator Smad 4 in NSCs or intraventicular infusion of noggin resulted in decreased neurogenesis and an increase in oligodendroglial progenitors migrating to the corpus callosum (Colak et al., (2008)).
Shh is a morphogen known to regulate neurogenesis and gliogenesis during development. It has also been demonstrated to increase granule cell precursor proliferation in the hippocampus (Wechsler-Reya and Scott, (1999)) as well as to increase stem/precursor cell proliferation in the SEZ (Palma et al., 2004). Two more recent studies showed that both NSCs and TaPs are responsive to Shh signalling (Ahn and Joyner, (2005)) and that Shh is essential for their maintenance (Balordi and Fishell, (2007)). In addition, genetic deletion of smoothened (Smo), a component of Shh signalling pathways, resulted in significant depletion of SGZ neurogenesis, while Smo overexpression led to upregulation of proliferation (Han et al., (2008)). Another easily diffusible molecule which has a variety of regulatory roles in the CNS is nitric oxide (NO). Interestingly, it appears that none of the major cell types in the rodent SEZ express molecules of the NO signalling pathway. However, processes of nitrergic neurons intercalating with neuroblasts at the lateral region of the SEZ have been shown to express neuronal NO synthase (Moreno-Lopez et al., (2004)). This has lead to the hypothesis that NO originating from neuronal processes might play a role in the SEZ. In keeping with this, inhibitors of NO signalling increase cell proliferation and NO synthase deficient mice also exhibit higher levels of proliferation in the SEZ (Moreno-Lopez et al., (2004); Packer et al., (2003); Pinnock et al., (2007)).
2.4.2. Cell to cell and cell to ECM interactions
Cell-cell and cell-ECM signalling could also regulate NSC and progenitor behaviour within adult niches. Members of the Notch signalling pathway (Notch and Jagged) are present in both the SEZ and SGZ as revealed by mRNA analysis (Stump et al., (2002)). In addition, ephrins and their receptors are present in adult neurogenic niches and both positively (Conover et al., (2000); ephrins-B2/3, Eph B1-3, A4) and negatively (Holmberg et al., (2005); ephrin-A2, Eph A7) regulate NSC proliferation. Intercellular interactions can also be mediated by cadherin-dependent adherens junctions or gap junctions formed by connexins. While there is evidence for both these interactions in other adult stem cell niches and in the embryonic NSC microenvironment (Lathia et al., (3267)), their functional relevance in adult NSC niches is yet to be determined. The role of the ECM molecules that are expressed in the adult neurogenic niches is also still elusive. Tenascin-C, a glycoprotein that has been shown to regulate growth factor activity during brain development (Garcion et al., (2004)) and that is highly expressed in the SEZ was shown to be dispensable for the neurogenic process (de Chevigny et al., (2006); Kazanis et al., (2007)). In a recent study, Kerever et al. (2006) demonstrated that the laminin-rich fractones can capture FGF2 and thus can regulate growth factor concentrations and activity in the SEZ, as has been suggested for other brain ECM molecules (Bandtlow and Zimmermann, (2000)). The expression analysis of chondroitin sulfate glycosaminoglycans (Sirko et al., (2007)) and of multiple chondroitin/dermatan sulfotransferases (Akita et al., (2008)) is also suggestive of a role of ECM in NSC/progenitor behavior, but direct evidence is still lacking.
2.4.3. Neurotransmission
Another class of molecules which seem to play a significant role in adult NSC niches is neurotransmitters. A series of recent studies have provided evidence for such an activity for γ-amino-butyric acid (GABA) in the SEZ and serotonin (5-HT) in the SGZ. GABA is the principle inhibitory neurotransmitter in the adult CNS but has an excitatory action in the SEZ (as during development) and the SGZ (Ge et al., (2007); Wang et al., (2003); Wang et al., (2005)). Isolated rat neuroblasts were shown to express the GABA-A receptor and GABA has been found to decrease neuroblast migration (Bolteus and Bordey, (2004)) and to cause cell cycle exit (Overstreet Wadiche et al., (2005)). Therefore, it has been suggested that it could participate in a feedback loop between NSCs and neuroblasts, controlling the number of neuroblasts produced at a given time (Liu et al., (2005)). Serotonin (5-HT), a neurotransmitter that is a therapeutic target in cases of depression, has recently been investigated for its role in modulating NSC behavior in the hippocampus (Santarelli et al., (2003)). Early studies depleting 5-HT in prenatal stages showed a reduction in cell proliferation in the SGZ, as well as in the SEZ (Brezun and Daszuta, (1999)). Furthermore, pharmacological studies have revealed the role of several 5-HT receptors on proliferation in the SEZ and SGZ. In the SEZ, proliferation is positively regulated by 5HT-1A and 2C receptors and negatively regulated by 5HT-1B and 2A/2C while in the SGZ it is positively regulated by 5HT-1A and negatively regulated by 5-HT2A/2C (Banasr et al., (2004); Radley and Jacobs, (2002)). The cell types which are responsive to serotonin are largely unknown; however a recent study showed that fluoxetine, a specific serotonin re-uptake inhibitor commonly used to treat depression, selectively affected the proliferation of immature neuroblasts (Encinas et al., (2006)). Nevertheless, the role of neurogenesis either as a cause of depression or as a target for therapeutic intervention is currently under intense investigation (Elder et al., (2006); Grote and Hannan, (2007)).
3. The NSC microenvironment in disease
Until recently the existence of adult neurogenesis leading to replacement and/or repair in the CNS was not widely accepted, because it was believed that no new cells could be added to the brain without affecting its function and because clinical experience suggested that cell loss in the CNS could not be repaired. Our current knowledge on the existence of small scale neurogenesis in the adult brain has not significantly altered these basic concepts, with two exceptions. Firstly, it is now established that the restricted turnover of neurons in the olfactory bulbs and, more importantly, in the hippocampus serves the plastic requirements of these systems. Second, it has been shown that the SEZ can contribute neuroblasts after injury or stroke, without being able to elicit adequate repair. However, the discovery of adult neurogenesis brings forward questions about the way this process might be linked with brain pathologies: is neurogenesis altered in cases of brain disease and is it a cause or a consequence of such diseases? In this section, therefore, the focus will be on existing data regarding the NSC microenvironment in different cases of brain disease.
3.1. Does altered neurogenesis contribute to brain pathologies?
Developmental malformations such as a small (microcephaly) or a large (hemimegalencephaly) brain, or focal abnormalities (focal cortical dysplasia) have been attributed to disturbed production of neurons and/or glial cells during embryonic development and are commonly associated with mental retardation and epilepsy (Pang et al., (2008)). The genes that have been correlated with these pathologies are associated with the intrinsic cell-cycle regulation rather than with the extracellular microenvironment (Bond and Woods, (2006)). Mutations in four genes have been identified for autosomal recessive primary microcephaly, a neurodevelomental disorder characterized by the congenital occurrence of a small brain with normal cytoarchitecture and no progressive cognitive decline and seizures (Woods et al., (2005)): i) Microcephalin, a gene encoding a protein that plays a role in controlling cell-cycle timing; ii) the abnormal spindle-like, microcephaly associated (ASPM) gene, that encodes a protein important for the formation of the central mitotic spindle; iii) Cyclin dependent kinase 5 regulatory associated protein 2 (CDK5RAP2) gene, encoding a centrosomal protein that interacts with gamma-tubulin ring complexes during spindle formation and iv) Centromere associated protein J (CENPJ) gene, encoding another centrosomal protein, important in microtubule nucleation and polymerization.
On the other hand, cortical malformations that are attributed to disturbed migration (either hypo or hyper migration) have been linked to genes that encode components of the extracellular environment or molecules that control the cellular response to that environment. One of the genes associated with the ectopic occurrence of neuroglial clusters at the ventricular surface (periventricular heterotopia), caused by the inability of newborn cells to migrate away from the ventricles, is Filamin A (FLNA; Lu et al., (2006)). FLNA is highly expressed in the neuroepithelium adjacent to the ventricles and is thought to contribute to the neuroepithelial layer structural integrity. The importance of this integrity in the correct migration of cells has also been demonstrated during development in mice deficient in N-cadherin signalling (Radice et al., (1997)) and in adults in mice with disrupted ependymal cell layer (Del Carmen Gomez-Roldan et al., (2008); Jimenez et al., (2001)), that are also characterized by the formation of cerebral heterotopias. In addition, genes that encode components of the microenvironment that surrounds neuronal progenitor basal processes (the basal membrane at the pial surface) have been correlated with loss of the gyri and sulci of the brain (classic and cobblestone lissencephaly; Pang et al., (2008)). In classic (type-I) lissencephaly the normal 6-layer structure of the cortex is lost. Absence of reelin (Bonneau et al., (2002)), a signalling glycoprotein secreted by early born neurons at the surface of the cortex, has been associated with this type of lissencephaly although most human cases have been correlated with genes encoding for proteins regulating microtubule assembly, such as lissencephaly 1 (LIS1; Reiner et al., (1993)), Doublecortin (Dcx; Gleeson et al., (1998); Pilz et al., (1998)) and tubulin alpha 1A (TUBA1A; Keays et al., (2007)). In cobblestone (type-II) lissencephaly cortical layers are disorganised as neurons over-migrate through the pial surface into the meninges to form ectopias. In animal models and humans, depletion of molecules of the pial basal membrane, of their receptors as well as of associated signalling components such as presenilin 1 (Hartmann et al., 1998), alpha6 integrin and integrin-linked kinase (Georges-Labouesse et al., (1998); Niewmierzycka et al., (2005)), result in phenotypes resembling this form of lissencephaly. In humans, cobblestone lissencephaly has been correlated with four genes: protein-O-mannosyltransferase 1 and 2 (POMT1-2), protein-O-mannose 1,2-N-acetylglucosaminyl-transferase (POMGnT1; Yoshida et al., (2001)) and Fukutin (Toda et al., (1994)). All these genes are involved in glycosylation of α-dystroglycan, a receptor for multiple ECM molecules. Mutations of the above-mentioned genes lead to compromised integrity of the basal lamina barrier at the pial surface, thus allowing hypermigration of cells (Guerrini and Marini, (2006); Pang et al., (2008)). Finally, alterations in the composition of the basement membrane molecules have been associated with brain malformations characterizing congenital muscular dystrophy (Philpot et al., (2000); Colognato et al., (2005)).
Although it is easy to identify altered neurogenesis and/or migration as the main cause of developmental brain malformations, it is very difficult to directly address whether defects in adult neurogenesis can also cause pathologies. This is due to the fact that samples from patients are taken only after a disease has been diagnosed, thus making it difficult to distinguish cause from effect. Also, genetically interfering with neurogenesis often results in perturbation of early development because, as illustrated in Figure 3, many of the regulatory mechanisms are shared in the embryo and the adult. Experiments where neurogenesis is genetically disturbed specifically in the postnatal brain are lacking and exogenously induced perturbations (irradiation, growth factor injections) have been studied only for short-term effects. One interesting study was that of Shors et al. ((2001)) in which abolishment of hippocampal neurogenesis was shown to result in impaired formation of trace memories. Therefore, the hypothesis that impaired adult neurogenesis can lead to brain pathology is based on indirect evidence, with the age-related decline in neurogenesis (Luo et al., (2006)) correlating with the occurrence of dementia and the demonstration that drug-induced upregulation of endogenous neurogenesis can be beneficial for the treatment of depression or dementia (Santarelli et al., (2003)).
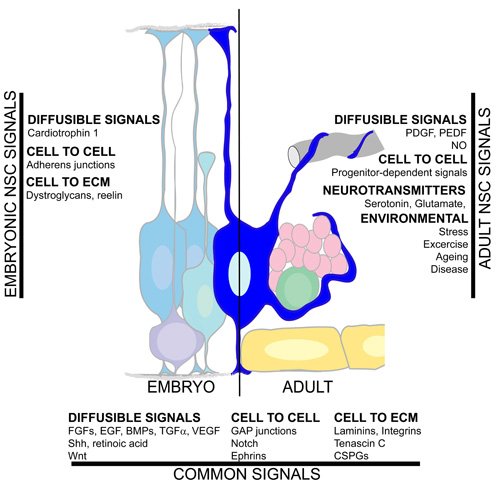
Figure 3. Summary of the signalling pathways in the neural stem cell microenvironment.
3.2. Is the adult neurogenic niche microenvironment altered in cases of brain disease?
Normal adult neurogenesis results in a limited volume of newly produced and functionally integrated cells and serves to maintain tissue homeostasis in only very specific systems; it might therefore be expected to be rather insensitive to stimuli derived outside of these systems. Surprisingly, however, there is accumulating evidence suggesting that both neurogenic niches are responsive to local signals generated from proximal tissue damage and also to more global signals from degeneration in remote areas and even as a result of changes in the external macro-environment of the organism (in the absence of tissue damage). For example, neurogenesis is consistently found to be increased in experimental models of ischemia/stroke in rodents both in the SEZ (Zhang et al., (2004); Zhang et al., (2008)) and the SGZ (Arvidsson et al., (2001); Bendel et al., (2005); Nakatomi et al., (2002)), as well as in human cases of stroke (Curtis et al., (2007)). Enhanced proliferation in the SEZ has also been reported in patients suffering from epileptic seizures (Grote and Hannan, (2007)) and multiple sclerosis (Nait-Oumesmar et al., (2007)). However, findings from animal experimental models that show recruitment of SEZ-generated oligodendrocytes in demyelination lesions (Nait-Oumesmar et al., (2007); Picard-Riera et al., (2002)) have not yet been confirmed in human tissue. Neurogenesis is increased in human cases and animal models of Huntington's disease while it is decreased in patients and animal models of Alzheimer's and Parkinson's disease (Curtis et al., (2007); Elder et al., (2006)). Finally, neurogenesis is generally reduced in cases of mood disorders, such as depression and stress (Elder et al., (2006); Grote and Hannan, (2007)).
How do these pathologies alter the environment of the SEZ and SGZ in such a way that NSC behaviour is altered? In the case of the SEZ, a global cue might be provided by the contact of the niche with the CSF, allowing it to sense information on the state of the organism as depicted by the concentrations of hormones and growth factors. Important information might also be present in the meninges-vessels-fractones continuum (Mercier et al., (2002)) and the astroglial syncytia (Giaume and Venance, (1998)). The SGZ, on the other hand, is incorporated within the hippocampal formation, a structure that participates in numerous neuronal networks, important in the processing of almost all of the externally acquired information and the formation of short-term, working memory. Turning to more local signals, little information exists regarding the neurogenic niche microenvironment in the above-mentioned neuropathologies. Ischemia results in the expansion of the SEZ with a parallel induction of hypoxic conditions and a transient decrease in vascular density (Thored et al., (2007)). The expanded SEZ is characterized by the presence of tenascin-C (Thored et al., (2007)), an ECM component normally expressed in this region (de Chevigny et al., (2006); Kazanis et al., (2007); Thomas et al., (1996)) and evidence from a recent microarray study showed increased expression of growth factors, morphogens and ECM molecules after stroke (Liu et al., (2007)). Similar upregulation of growth factors, neurotrophic and angiogenic factors and ECM restructuring have been reported after experimentally-induced seizures in the hippocampus (Liu et al., (2007); Newton et al., (2003)), another condition of upregulated neurogenesis. Finally, an interesting finding is that increased neurogenesis is observed only in excitotoxic animal models of Huntington's disease while no change is observed in transgenic models, in which there is minimal cell loss. Therefore, it emerges that the presence of inflammatory signals in the SEZ might be an important factor in the regulation of neurogenesis (Phillips et al., (2005)), just as inflammation has been proposed to promote remyelination following demyelination, perhaps the best example of repair in the mammalian CNS (Zawadzka and Franklin, (2007)).
Acknowledgements
We are grateful to Dr. Veronique Marthiens for her instructive comments. The authors are funded by grants of the NIH (National Institute of Biomedical Imaging and Bioengineering Quantum Grant Project 1P20EB00706), the Biotechnology and Biological Sciences Research Council (BBSRC), the Medical Research Council (MRC) and the NIH- Cambridge Graduate Partnership Program.
Glossary
brain-derived neurotrophic factor;
bone morphogenetic protein;
central nervous system;
cerebrospinal fluid;
extracellular matrix;
fibroblast growth factor;
leukaemia inhibitory factor;
neuroblast;
neuroepithelial cell;
neural stem cell;
platelet-derived growth factor;
pigment epithelium-derived factor;
radial glial cell;
subependymal zone;
subgranular zone;
subventricular zone;
transit amplifying precursor;
transforming growth factor;
vascular endothelial growth factor;
ventricular zone.
Copyright: © 2008 Ilias Kazanis, Justin Lathia, Lara Moss, and Charles ffrench-Constant.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
§ To whom correspondence should be addressed. E-mail: ik255@cam.ac.uk .
* Edited by: David Scadden. Last revised June 16, 2008. Published August 31, 2008. This chapter should be cited as: Kazanis, I., Lathia, J., Moss, L., and ffrench-Constant, C., The neural stem cell microenvironment (August 31, 2008), StemBook, ed. The Stem Cell Research Community, StemBook, doi/10.3824/stembook.1.15.1, https://www.stembook.org .
References
- Aaku-Saraste, E. Hellwig, A. Huttner, W. B. (1996). Loss of occludin and functional tight junctions, but not ZO-1, during neural tube closure–remodeling of the neuroepithelium prior to neurogenesis. Dev Biol 180, 664–679. Abstract Abstract
- Ahn, S. Joyner, A. L. (2005). In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature 437, 894–897. Abstract Abstract
- Akita, K. von Holst, A. Furukawa, Y. Mikami, T. Sugahara, K. Faissner, A. (2008). Expression of multiple chondroitin/dermatan sulfotransferases in the neurogenic regions of the embryonic and adult central nervous system implies that complex chondroitin sulfates have a role in neural stem cell maintenance. Stem Cells 26, 798–809. Abstract Abstract
- Alvarez-Buylla, A. Garcia-Verdugo, J. M. Tramontin, A. D. (2001). A unified hypothesis on the lineage of neural stem cells. Nat Rev Neurosci 2, 287–293. Abstract Abstract
- Androutsellis-Theotokis, A. Leker, R. R. Soldner, F. Hoeppner, D. J. Ravin, R. Poser, S. W. Rueger, M. A. Bae, S. K. Kittappa, R. McKay, R. D. (2006). Notch signalling regulates stem cell numbers in vitro and in vivo. Nature 442, 823–826. Abstract Abstract
- Arvidsson, A. Kokaia, Z. Lindvall, O. (2001). N-methyl-D-aspartate receptor-mediated increase of neurogenesis in adult rat dentate gyrus following stroke. Eur J Neurosci 14, 10–18. Abstract Abstract
- Assimacopoulos, S. Grove, E. A. Ragsdale, C. W. (2003). Identification of a Pax6-dependent epidermal growth factor family signalling source at the lateral edge of the embryonic cerebral cortex. J Neurosci 23, 6399–6403. Abstract Abstract
- Backman, M. Machon, O. Mygland, L. van den Bout, C. J. Zhong, W. Taketo, M. M. Krauss, S. (2005). Effects of canonical Wnt signalling on dorso-ventral specification of the mouse telencephalon. Dev Biol 279, 155–168. Abstract Abstract
- Ballas, N. Mandel, G. (2005). The many faces of REST oversee epigenetic programming of neuronal genes. Curr Opin Neurobiol 15, 500–506. Abstract Abstract
- Balordi, F. Fishell, G. (2007). Hedgehog signalling in the subventricular zone is required for both the maintenance of stem cells and the migration of newborn neurons. J Neurosci 27, 5936–5947. Abstract Abstract
- Banasr, M. Hery, M. Printemps, R. Daszuta, A. (2004). Serotonin-induced increases in adult cell proliferation and neurogenesis are mediated through different and common 5-HT receptor subtypes in the dentate gyrus and the subventricular zone. Neuropsychopharmacology 29, 450–460. Abstract Abstract
- Bandtlow, C. E. Zimmermann, D. R. (2000). Proteoglycans in the developing brain: new conceptual insights for old proteins. Physiol Rev 80, 1267–1290. Abstract Abstract
- Bansal, R. Lakhina, V. Remedios, R. Tole, S. (2003). Expression of FGF receptors 1, 2, 3 in the embryonic and postnatal mouse brain compared with Pdgfralpha, Olig2 and Plp/dm20: implications for oligodendrocyte development. Dev Neurosci 25, 83–95. Abstract Abstract
- Barnabe-Heider, F. Wasylnka, J. A. Fernandes, K. J. Porsche, C. Sendtner, M. Kaplan, D. R. Miller, F. D. (2005). Evidence that embryonic neurons regulate the onset of cortical gliogenesis via cardiotrophin-1. Neuron 48, 253–265. Abstract Abstract
- Beckers, J. Caron, A. Hrabe de Angelis, M. Hans, S. Campos-Ortega, J. A. Gossler, A. (2000). Distinct regulatory elements direct delta1 expression in the nervous system and paraxial mesoderm of transgenic mice. Mech Dev 95, 23–34. Abstract Abstract
- Becq, H. Jorquera, I. Ben-Ari, Y. Weiss, S. Represa, A. (2005). Differential properties of dentate gyrus and CA1 neural precursors. J Neurobiol 62, 243–261. Abstract Abstract
- Beggs, H. E. Schahin-Reed, D. Zang, K. Goebbels, S. Nave, K. A. Gorski, J. Jones, K. R. Sretavan, D. Reichardt, L. F. (2003). FAK deficiency in cells contributing to the basal lamina results in cortical abnormalities resembling congenital muscular dystrophies. Neuron 40, 501–514. Abstract Abstract
- Bendel, O. Bueters, T. von Euler, M. Ove Ogren, S. Sandin, J. von Euler, G. (2005). Reappearance of hippocampal CA1 neurons after ischemia is associated with recovery of learning and memory. J Cereb Blood Flow Metab 25, 1586–1595. Abstract Abstract
- Bertrand, N. Castro, D. S. Guillemot, F. (2002). Proneural genes and the specification of neural cell types. Nat Rev Neurosci 3, 517–530. Abstract Abstract
- Bertrand, N. Dahmane, N. (2006). Sonic hedgehog signalling in forebrain development and its interactions with pathways that modify its effects. Trends Cell Biol 16, 597–605. Abstract Abstract
- Biron, V. L. McManus, K. J. Hu, N. Hendzel, M. J. Underhill, D. A. (2004). Distinct dynamics and distribution of histone methyl-lysine derivatives in mouse development. Dev Biol 276, 337–351. Abstract Abstract
- Bittman, K. S. LoTurco, J. J. (1999). Differential regulation of connexin 26 and 43 in murine neocortical precursors. Cereb Cortex 9, 188–195. Abstract Abstract
- Bolteus, A. J. Bordey, A. (2004). GABA release and uptake regulate neuronal precursor migration in the postnatal subventricular zone. J Neurosci 24, 7623–7631. Abstract Abstract
- Bond, J. Woods, C. G. (2006). Cytoskeletal genes regulating brain size. Curr Opin Cell Biol 18, 95–101. Abstract Abstract
- Bonneau, D. Toutain, A. Laquerriere, A. Marret, S. Saugier-Veber, P. Barthez, M. A. Radi, S. Biran-Mucignat, V. Rodriguez, D. Gelot, A. (2002). X-linked lissencephaly with absent corpus callosum and ambiguous genitalia (XLAG): clinical, magnetic resonance imaging, and neuropathological findings. Ann Neurol 51, 340–349. Abstract Abstract
- Brannvall, K. Korhonen, L. Lindholm, D. (2002). Estrogen-receptor-dependent regulation of neural stem cell proliferation and differentiation. Mol Cell Neurosci 21, 512–520. Abstract Abstract
- Brezun, J. M. Daszuta, A. (1999). Depletion in serotonin decreases neurogenesis in the dentate gyrus and the subventricular zone of adult rats. Neuroscience 89, 999–1002. Abstract Abstract
- Bruni, J. E. (1998). Ependymal development, proliferation, and functions: a review. Microsc Res Tech 41, 2–13. Abstract Abstract
- Bull, N. D. Bartlett, P. F. (2005). The adult mouse hippocampal progenitor is neurogenic but not a stem cell. J Neurosci 25, 10815–10821. Abstract Abstract
- Campbell, K. (2003). Dorsal-ventral patterning in the mammalian telencephalon. Curr Opin Neurobiol 13, 50–56. Abstract Abstract
- Campos, L. S. Leone, D. P. Relvas, J. B. Brakebusch, C. Fassler, R. Suter, U. ffrench-Constant, C. (2004). Beta1 integrins activate a MAPK signalling pathway in neural stem cells that contributes to their maintenance. Development 131, 3433–3444. Abstract Abstract
- Cao, L. Jiao, X. Zuzga, D. S. Liu, Y. Fong, D. M. Young, D. During, M. J. (2004). VEGF links hippocampal activity with neurogenesis, learning and memory. Nat Genet 36, 827–835. Abstract Abstract
- Cao, X. Yeo, G. Muotri, A. R. Kuwabara, T. Gage, F. H. (2006). Noncoding RNAs in the mammalian central nervous system. Annu Rev Neurosci 29, 77–103. Abstract Abstract
- Capela, A. Temple, S. (2002). LeX/ssea-1 is expressed by adult mouse CNS stem cells, identifying them as nonependymal. Neuron 35, 865–875. Abstract Abstract
- Carmeliet, P. (2003). Angiogenesis in health and disease. Nat Med 9, 653–660. Abstract Abstract
- Chen, D. McKearin, D. (2005). Gene circuitry controlling a stem cell niche. Curr Biol 15, 179–184. Abstract Abstract
- Cheng, S. Shakespeare, T. Mui, R. White, T. W. Valdimarsson, G. (2004). Connexin 48.5 is required for normal cardiovascular function and lens development in zebrafish embryos. J Biol Chem 279, 36993–37003. Abstract Abstract
- Chenn, A. Walsh, C. A. (2002). Regulation of cerebral cortical size by control of cell cycle exit in neural precursors. Science 297, 365–369. Abstract Abstract
- Chi, C. L. Martinez, S. Wurst, W. Martin, G. R. (2003). The isthmic organizer signal FGF8 is required for cell survival in the prospective midbrain and cerebellum. Development 130, 2633–2644. Abstract Abstract
- Chiang, C. Litingtung, Y. Lee, E. Young, K. E. Corden, J. L. Westphal, H. Beachy, P. A. (1996). Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 383, 407–413. Abstract Abstract
- Colak, D. Mori, T. Brill, M. S. Pfeifer, A. Falk, S. Deng, C. Monteiro, R. Mummery, C. Sommer, L. Gotz, M. (2008). Adult neurogenesis requires Smad4-mediated bone morphogenic protein signalling in stem cells. J Neurosci 28, 434–446. Abstract Abstract
- Colognato, H. ffrench-Constant, C. Feltri, M. L. (2005). Human diseases reveal novel roles for neural laminins. Trends Neurosci 28, 480–486. Abstract Abstract
- Conover, J. C. Doetsch, F. Garcia-Verdugo, J. M. Gale, N. W. Yancopoulos, G. D. Alvarez-Buylla, A. (2000). Disruption of Eph/ephrin signalling affects migration and proliferation in the adult subventricular zone. Nat Neurosci 3, 1091–1097. Abstract Abstract
- Corbit, K. C. Aanstad, P. Singla, V. Norman, A. R. Stainier, D. Y. Reiter, J. F. (2005). Vertebrate Smoothened functions at the primary cilium. Nature 437, 1018–1021. Abstract Abstract
- Craig, C. G. Tropepe, V. Morshead, C. M. Reynolds, B. A. Weiss, S. van der Kooy, D. (1996). In vivo growth factor expansion of endogenous subependymal neural precursor cell populations in the adult mouse brain. J Neurosci 16, 2649–2658. Abstract Abstract
- Curtis, M. A. Eriksson, P. S. Faull, R. L. (2007). Progenitor cells and adult neurogenesis in neurodegenerative diseases and injuries of the basal ganglia. Clin Exp Pharmacol Physiol 34, 528–532. Abstract Abstract
- Dahmane, N. Sanchez, P. Gitton, Y. Palma, V. Sun, T. Beyna, M. Weiner, H. Ruiz i Altaba, A. (2001). The Sonic Hedgehog-Gli pathway regulates dorsal brain growth and tumorigenesis. Development 128, 5201–5212. Abstract Abstract
- Dang, L. Yoon, K. Wang, M. Gaiano, N. (2006). Notch3 signalling promotes radial glial/progenitor character in the mammalian telencephalon. Dev Neurosci 28, 58–69. Abstract Abstract
- Darsalia, V. Kallur, T. Kokaia, Z. (2007). Survival, migration and neuronal differentiation of human fetal striatal and cortical neural stem cells grafted in stroke-damaged rat striatum. Eur J Neurosci 26, 605–614. Abstract Abstract
- Del Carmen Gomez-Roldan, M. Perez-Martin, M. Capilla-Gonzalez, V. Cifuentes, M. Perez, J. Garcia-Verdugo, J. M. Fernandez-Llebrez, P. (2008). Neuroblast proliferation on the surface of the adult rat striatal wall after focal ependymal loss by intracerebroventricular injection of neuraminidase. J Comp Neurol 507, 1571–1587. Abstract Abstract
- Depaepe, V. Suarez-Gonzalez, N. Dufour, A. Passante, L. Gorski, J. A. Jones, K. R. Ledent, C. Vanderhaeghen, P. (2005). Ephrin signalling controls brain size by regulating apoptosis of neural progenitors. Nature 435, 1244–1250. Abstract Abstract
- Dessaud, E. Yang, L. L. Hill, K. Cox, B. Ulloa, F. Ribeiro, A. Mynett, A. Novitch, B. G. Briscoe, J. (2007). Interpretation of the sonic hedgehog morphogen gradient by a temporal adaptation mechanism. Nature 450, 717–720. Abstract Abstract
- Doetsch, F. Garcia-Verdugo, J. M. Alvarez-Buylla, A. (1997). Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J Neurosci 17, 5046–5061. Abstract Abstract
- Doetsch, F. Garcia-Verdugo, J. M. Alvarez-Buylla, A. (1999). Regeneration of a germinal layer in the adult mammalian brain. Proc Natl Acad Sci USA 96, 11619–11624. Abstract Abstract
- Doetsch, F. Petreanu, L. Caille, I. Garcia-Verdugo, J. M. Alvarez-Buylla, A. (2002). EGF converts transit-amplifying neurogenic precursors in the adult brain into multipotent stem cells. Neuron 36, 1021–1034. Abstract Abstract
- Elder, G. A. De Gasperi, R. Gama Sosa, M. A. (2006). Research update: neurogenesis in adult brain and neuropsychiatric disorders. Mt Sinai J Med 73, 931–940. Abstract Abstract
- Elias, L. A. Wang, D. D. Kriegstein, A. R. (2007). Gap junction adhesion is necessary for radial migration in the neocortex. Nature 448, 901–907. Abstract Abstract
- Encinas, J. M. Vaahtokari, A. Enikolopov, G. (2006). Fluoxetine targets early progenitor cells in the adult brain. Proc Natl Acad Sci USA 103, 8233–8238. Abstract Abstract
- Episkopou, V. (2005). SOX2 functions in adult neural stem cells. Trends Neurosci 28, 219–221. Abstract Abstract
- Fuller, M. T. Spradling, A. C. (2007). Male and female Drosophila germline stem cells: two versions of immortality. Science 316, 402–404. Abstract Abstract
- Fushiki, S. Perez Velazquez, J. L. Zhang, L. Bechberger, J. F. Carlen, P. L. Naus, C. C. (2003). Changes in neuronal migration in neocortex of connexin43 null mutant mice. J Neuropathol Exp Neurol 62, 304–314. Abstract Abstract
- Gaiano, N. Nye, J. S. Fishell, G. (2000). Radial glial identity is promoted by Notch1 signalling in the murine forebrain. Neuron 26, 395–404. Abstract Abstract
- Gal, J. S. Morozov, Y. M. Ayoub, A. E. Chatterjee, M. Rakic, P. Haydar, T. F. (2006). Molecular and morphological heterogeneity of neural precursors in the mouse neocortical proliferative zones. J Neurosci 26, 1045–1056. Abstract Abstract
- Garcion, E. Halilagic, A. Faissner, A. ffrench-Constant, C. (2004). Generation of an environmental niche for neural stem cell development by the extracellular matrix molecule tenascin C. Development 131, 3423–3432. Abstract Abstract
- Gato, A. Moro, J. A. Alonso, M. I. Bueno, D. De La Mano, A. Martin, C. (2005). Embryonic cerebrospinal fluid regulates neuroepithelial survival, proliferation, and neurogenesis in chick embryos. Anat Rec A Discov Mol Cell Evol Biol 284, 475–484. Abstract Abstract
- Ge, S. Pradhan, D. A. Ming, G. L. Song, H. (2007). GABA sets the tempo for activity-dependent adult neurogenesis. Trends Neurosci 30, 1–8. Abstract Abstract
- Georges-Labouesse, E. Mark, M. Messaddeq, N. Gansmuller, A. (1998). Essential role of alpha 6 integrins in cortical and retinal lamination. Curr Biol 8, 983–986. Abstract Abstract
- Giaume, C. Venance, L. (1998). Intercellular calcium signalling and gap junctional communication in astrocytes. Glia 24, 50–64. Abstract Abstract
- Gleeson, J. G. Allen, K. M. Fox, J. W. Lamperti, E. D. Berkovic, S. Scheffer, I. Cooper, E. C. Dobyns, W. B. Minnerath, S. R. Ross, M.E. (1998). Doublecortin, a brain-specific gene mutated in human X-linked lissencephaly and double cortex syndrome, encodes a putative signalling protein. Cell 92, 63–72. Abstract Abstract
- Goldman, S. A. Nottebohm, F. (1983). Neuronal production, migration, and differentiation in a vocal control nucleus of the adult female canary brain. Proc Natl Acad Sci USA 80, 2390–2394. Abstract Abstract
- Gotz, M. Huttner, W. B. (2005). The cell biology of neurogenesis. Nat Rev Mol Cell Biol 6, 777–788. Abstract Abstract
- Graus-Porta, D. Blaess, S. Senften, M. Littlewood-Evans, A. Damsky, C. Huang, Z. Orban, P. Klein, R. Schittny, J. C. Muller, U. (2001). Beta1-class integrins regulate the development of laminae and folia in the cerebral and cerebellar cortex. Neuron 31, 367–379. Abstract Abstract
- Greferath, U. Canty, A. J. Messenger, J. Murphy, M. (2002). Developmental expression of EphA4-tyrosine kinase receptor in the mouse brain and spinal cord. Gene Expr Patterns 2, 267–274. Abstract Abstract
- Grote, H. E. Hannan, A. J. (2007). Regulators of adult neurogenesis in the healthy and diseased brain. Clin Exp Pharmacol Physiol 34, 533–545. Abstract Abstract
- Guerrini, R. Marini, C. (2006). Genetic malformations of cortical development. Exp Brain Res 173, 322–333.
- Guillemot, F. (2005). Cellular and molecular control of neurogenesis in the mammalian telencephalon. Curr Opin Cell Biol 17, 639–647. Abstract Abstract
- Gunhaga, L. Marklund, M. Sjodal, M. Hsieh, J. C. Jessell, T. M. Edlund, T. (2003). Specification of dorsal telencephalic character by sequential Wnt and FGF signalling. Nat Neurosci 6, 701–707. Abstract Abstract
- Hall, P. E. Lathia, J. D. Miller, N. G. Caldwell, M. A. ffrench-Constant, C. (2006). Integrins are markers of human neural stem cells. Stem Cells 24, 2078–2084. Abstract Abstract
- Han, Y. G. Spassky, N. Romaguera-Ros, M. Garcia-Verdugo, J. M. Aguilar, A. Schneider-Maunoury, S. Alvarez-Buylla, A. (2008). Hedgehog signalling and primary cilia are required for the formation of adult neural stem cells. Nat Neurosci 11, 277–284. Abstract Abstract
- Hartfuss, E. Galli, R. Heins, N. Gotz, M. (2001). Characterization of CNS precursor subtypes and radial glia. Dev Biol 229, 15–30. Abstract Abstract
- Hartfuss, E. Forster, E. Bock, H. H. Hack, M. A. Leprince, P. Luque, J. M. Herz, J. Frotscher, M. Gotz, M. (2003). Reelin signalling directly affects radial glia morphology and biochemical maturation. Development 130, 4597–4609. Abstract Abstract
- Haubst, N. Georges-Labouesse, E. De Arcangelis, A. Mayer, U. Gotz, M. (2006). Basement membrane attachment is dispensable for radial glial cell fate and for proliferation, but affects positioning of neuronal subtypes. Development 133, 3245–3254. Abstract Abstract
- Herken, R. Gotz, W. Wattjes, K. H. (1989). Initial development of capillaries in the neuroepithelium of the mouse. J Anat 164, 85–92. Abstract Abstract
- Hermanson, O. Glass, C. K. Rosenfeld, M. G. (2002). Nuclear receptor coregulators: multiple modes of modification. Trends Endocrinol Metab 13, 55–60. Abstract Abstract
- Hirabayashi, Y. Itoh, Y. Tabata, H. Nakajima, K. Akiyama, T. Masuyama, N. Gotoh, Y. (2004). The Wnt/beta-catenin pathway directs neuronal differentiation of cortical neural precursor cells. Development 131, 2791–2801. Abstract Abstract
- Hitoshi, S. Alexson, T. Tropepe, V. Donoviel, D. Elia, A. J. Nye, J. S. Conlon, R. A. Mak, T. W. Bernstein, A. van der Kooy, D. (2002). Notch pathway molecules are essential for the maintenance, but not the generation, of mammalian neural stem cells. Genes Dev 16, 846–858. Abstract Abstract
- Holmberg, J. Armulik, A. Senti, K. A. Edoff, K. Spalding, K. Momma, S. Cassidy, R. Flanagan, J. G. Frisen, J. (2005). Ephrin-A2 reverse signalling negatively regulates neural progenitor proliferation and neurogenesis. Genes Dev 19, 462–471. Abstract Abstract
- Hui, C. C. Slusarski, D. Platt, K. A. Holmgren, R. Joyner, A. L. (1994). Expression of three mouse homologs of the Drosophila segment polarity gene cubitus interruptus, Gli, Gli-2, and Gli-3, in ectoderm- and mesoderm-derived tissues suggests multiple roles during postimplantation development. Dev Biol 162, 402–413. Abstract Abstract
- Hunter, D. D. Llinas, R. Ard, M. Merlie, J. P. Sanes, J. R. (1992). Expression of s-laminin and laminin in the developing rat central nervous system. J Comp Neurol 323, 238–251. Abstract Abstract
- Jackson, E. L. Garcia-Verdugo, J. M. Gil-Perotin, S. Roy, M. Quinones-Hinojosa, A. VandenBerg, S. Alvarez-Buylla, A. (2006). PDGFR alpha-positive B cells are neural stem cells in the adult SVZ that form glioma-like growths in response to increased PDGF signalling. Neuron 51, 187–199. Abstract Abstract
- Jaworski, D. M. Fager, N. (2000). Regulation of tissue inhibitor of metalloproteinase-3 (Timp-3) mRNA expression during rat CNS development. J Neurosci Res 61, 396–408. Abstract Abstract
- Jimenez, A. J. Tome, M. Paez, P. Wagner, C. Rodriguez, S. Fernandez-Llebrez, P. Rodriguez, E. M. Perez-Figares, J. M. (2001). A programmed ependymal denudation precedes congenital hydrocephalus in the hyh mutant mouse. J Neuropathol Exp Neurol 60, 1105–1119. Abstract Abstract
- Jin, K. Zhu, Y. Sun, Y. Mao, X. O. Xie, L. Greenberg, D. A. (2002). Vascular endothelial growth factor (VEGF) stimulates neurogenesis in vitro and in vivo. Proc Natl Acad Sci USA 99, 11946–11950. Abstract Abstract
- Kadowaki, M. Nakamura, S. Machon, O. Krauss, S. Radice, G. L. Takeichi, M. (2007). N-cadherin mediates cortical organization in the mouse brain. Dev Biol 304, 22–33. Abstract Abstract
- Kalyani, A. Hobson, K. Rao, M. S. (1997). Neuroepithelial stem cells from the embryonic spinal cord: isolation, characterization, and clonal analysis. Dev Biol 186, 202–223. Abstract Abstract
- Kalyani, A. J. Mujtaba, T. Rao, M. S. (1999). Expression of EGF receptor and FGF receptor isoforms during neuroepithelial stem cell differentiation. J Neurobiol 38, 207–224. Abstract Abstract
- Kazanis, I. Belhadi, A. Faissner, A. Ffrench-Constant, C. (2007). The adult mouse subependymal zone regenerates efficiently in the absence of tenascin-C. J Neurosci 27, 13991–13996. Abstract Abstract
- Keays, D. A. Tian, G. Poirier, K. Huang, G. J. Siebold, C. Cleak, J. Oliver, P. L. Fray, M. Harvey, R. J. Molnar, Z. (2007). Mutations in alpha-tubulin cause abnormal neuronal migration in mice and lissencephaly in humans. Cell 128, 45–57. Abstract Abstract
- Kerever, A. Schnack, J. Vellinga, D. Ichikawa, N. Moon, C. Arikawa-Hirasawa, E. Efird, J. T. Mercier, F. (2007). Novel extracellular matrix structures in the neural stem cell niche capture the neurogenic factor fibroblast growth factor 2 from the extracellular milieu. Stem Cells 25, 2146–2157. Abstract Abstract
- Kilpatrick, T. J. Talman, P. S. Bartlett, P. F. (1993). The differentiation and survival of murine neurons in vitro is promoted by soluble factors produced by an astrocytic cell line. J Neurosci Res 35, 147–161. Abstract Abstract
- Kim, A. S. Lowenstein, D. H. Pleasure, S. J. (2001). Wnt receptors and Wnt inhibitors are expressed in gradients in the developing telencephalon. Mech Dev 103, 167–172. Abstract Abstract
- Kosodo, Y. Roper, K. Haubensak, W. Marzesco, A. M. Corbeil, D. Huttner, W. B. (2004). Asymmetric distribution of the apical plasma membrane during neurogenic divisions of mammalian neuroepithelial cells. EMBO J 23, 2314–2324. Abstract Abstract
- Kostyszyn, B. Cowburn, R. F. Seiger, A. Kjaeldgaard, A. Sundstrom, E. (2004). Distribution of presenilin 1 and 2 and their relation to Notch receptors and ligands in human embryonic/foetal central nervous system. Brain Res Dev Brain Res 151, 75–86. Abstract Abstract
- Kuhn, H. G. Winkler, J. Kempermann, G. Thal, L. J. Gage, F. H. (1997). Epidermal growth factor and fibroblast growth factor-2 have different effects on neural progenitors in the adult rat brain. J Neurosci 17, 5820–5829. Abstract Abstract
- Lathia, J. D. Patton, B. Eckley, D. M. Magnus, T. Mughal, M. R. Sasaki, T. Caldwell, M. A. Rao, M. S. Mattson, M. P. ffrench-Constant, C. (2007a). Patterns of laminins and integrins in the embryonic ventricular zone of the CNS. J Comp Neurol 505, 630–643. Abstract Abstract
- Lathia, J. D. Rao, M. S. Mattson, M. P. Ffrench-Constant, C. (2007b). The microenvironment of the embryonic neural stem cell: lessons from adult niches?. Dev Dyn 236, 3267–3282. Abstract Abstract
- Lele, Z. Folchert, A. Concha, M. Rauch, G. J. Geisler, R. Rosa, F. Wilson, S. W. Hammerschmidt, M. Bally-Cuif, L. (2002). parachute/n-cadherin is required for morphogenesis and maintained integrity of the zebrafish neural tube. Development 129, 3281–3294. Abstract Abstract
- Leone, D. P. Relvas, J. B. Campos, L. S. Hemmi, S. Brakebusch, C. Fassler, R. Ffrench-Constant, C. Suter, U. (2005). Regulation of neural progenitor proliferation and survival by beta1 integrins. J Cell Sci 118, 2589–2599. Abstract Abstract
- Leventhal, C. Rafii, S. Rafii, D. Shahar, A. Goldman, S. A. (1999). Endothelial trophic support of neuronal production and recruitment from the adult mammalian subependyma. Mol Cell Neurosci 13, 450–464. Abstract Abstract
- Li, W. Cogswell, C. A. LoTurco, J. J. (1998). Neuronal differentiation of precursors in the neocortical ventricular zone is triggered by BMP. J Neurosci 18, 8853–8862. Abstract Abstract
- Li, W. LoTurco, J. J. (2000). Noggin is a negative regulator of neuronal differentiation in developing neocortex. Dev Neurosci 22, 68–73. Abstract Abstract
- Lim, D. A. Tramontin, A. D. Trevejo, J. M. Herrera, D. G. Garcia-Verdugo, J. M. Alvarez-Buylla, A. (2000). Noggin antagonizes BMP signalling to create a niche for adult neurogenesis. Neuron 28, 713–726. Abstract Abstract
- Liu, X. Wang, Q. Haydar, T. F. Bordey, A. (2005). Nonsynaptic GABA signalling in postnatal subventricular zone controls proliferation of GFAP-expressing progenitors. Nat Neurosci 8, 1179–1187. Abstract Abstract
- Liu, X. S. Zhang, Z. G. Zhang, R. L. Gregg, S. R. Meng, H. Chopp, M. (2007). Comparison of in vivo and in vitro gene expression profiles in subventricular zone neural progenitor cells from the adult mouse after middle cerebral artery occlusion. Neuroscience 146, 1053–1061. Abstract Abstract
- Lu, J. Tiao, G. Folkerth, R. Hecht, J. Walsh, C. Sheen, V. (2006). Overlapping expression of ARFGEF2 and Filamin A in the neuroependymal lining of the lateral ventricles: insights into the cause of periventricular heterotopia. J Comp Neurol 494, 476–484. Abstract Abstract
- Luo, J. Daniels, S. B. Lennington, J. B. Notti, R. Q. Conover, J. C. (2006). The aging neurogenic subventricular zone. Aging Cell 5, 139–152. Abstract Abstract
- Machon, O. van den Bout, C. J. Backman, M. Kemler, R. Krauss, S. (2003). Role of beta-catenin in the developing cortical and hippocampal neuroepithelium. Neuroscience 122, 129–143. Abstract Abstract
- Machon, O. Backman, M. Machonova, O. Kozmik, Z. Vacik, T. Andersen, L. Krauss, S. (2007). A dynamic gradient of Wnt signalling controls initiation of neurogenesis in the mammalian cortex and cellular specification in the hippocampus. Dev Biol 311, 223–237. Abstract Abstract
- Mackarehtschian, K. Lau, C. K. Caras, I. McConnell, S. K. (1999). Regional differences in the developing cerebral cortex revealed by ephrin-A5 expression. Cereb Cortex 9, 601–610. Abstract Abstract
- Maden, M. (2002). Retinoid signalling in the development of the central nervous system. Nat Rev Neurosci 3, 843–853. Abstract Abstract
- Malatesta, P. Hartfuss, E. Gotz, M. (2000). Isolation of radial glial cells by fluorescent-activated cell sorting reveals a neuronal lineage. Development 127, 5253–5263. Abstract Abstract
- Maric, D. Fiorio Pla, A. Chang, Y. H. Barker, J. L. (2007). Self-renewing and differentiating properties of cortical neural stem cells are selectively regulated by basic fibroblast growth factor (FGF) signalling via specific FGF receptors. J Neurosci 27, 1836–1852. Abstract Abstract
- Marshall, H. Nonchev, S. Sham, M. H. Muchamore, I. Lumsden, A. Krumlauf, R. (1992). Retinoic acid alters hindbrain Hox code and induces transformation of rhombomeres 2/3 into a 4/5 identity. Nature 360, 737–741. Abstract Abstract
- Martinez-Cerdeno, V. Noctor, S. C. Kriegstein, A. R. (2006). The role of intermediate progenitor cells in the evolutionary expansion of the cerebral cortex. Cereb Cortex 16(Suppl 1), i152–161. Abstract Abstract
- Marzesco, A. M. Janich, P. Wilsch-Brauninger, M. Dubreuil, V. Langenfeld, K. Corbeil, D. Huttner, W. B. (2005). Release of extracellular membrane particles carrying the stem cell marker prominin-1 (CD133) from neural progenitors and other epithelial cells. J Cell Sci 118, 2849–2858. Abstract Abstract
- Mashayekhi, F. Salehi, Z. (2006). The importance of cerebrospinal fluid on neural cell proliferation in developing chick cerebral cortex. Eur J Neurol 13, 266–272. Abstract Abstract
- Mason, I. (2007). Initiation to end point: the multiple roles of fibroblast growth factors in neural development. Nat Rev Neurosci 8, 583–596. Abstract Abstract
- Mercier, F. Kitasako, J. T. Hatton, G. I. (2002). Anatomy of the brain neurogenic zones revisited: fractones and the fibroblast/macrophage network. J Comp Neurol 451, 170–188. Abstract Abstract
- Mi, H. Haeberle, H. Barres, B. A. (2001). Induction of astrocyte differentiation by endothelial cells. J Neurosci 21, 1538–1547. Abstract Abstract
- Miyan, J. A. Zendah, M. Mashayekhi, F. Owen-Lynch, P. J. (2006). Cerebrospinal fluid supports viability and proliferation of cortical cells in vitro, mirroring in vivo development. Cerebrospinal Fluid Res 3, 2. Abstract Abstract
- Mizutani, K. Yoon, K. Dang, L. Tokunaga, A. Gaiano, N. (2007). Differential Notch signalling distinguishes neural stem cells from intermediate progenitors. Nature 449, 351–355. Abstract Abstract
- Moreno-Lopez, B. Romero-Grimaldi, C. Noval, J. A. Murillo-Carretero, M. Matarredona, E. R. Estrada, C. (2004). Nitric oxide is a physiological inhibitor of neurogenesis in the adult mouse subventricular zone and olfactory bulb. J Neurosci 24, 85–95. Abstract Abstract
- Muenke, M. Cohen, M. M. (2000). Genetic approaches to understanding brain development: holoprosencephaly as a model. Ment Retard Dev Disabil Res Rev 6, 15–21. Abstract Abstract
- Murphy, M. Drago, J. Bartlett, P. F. (1990). Fibroblast growth factor stimulates the proliferation and differentiation of neural precursor cells in vitro. J Neurosci Res 25, 463–475. Abstract Abstract
- Nait-Oumesmar, B. Picard-Riera, N. Kerninon, C. Decker, L. Seilhean, D. Hoglinger, G. U. Hirsch, E. C. Reynolds, R. Baron-Van Evercooren, A. (2007). Activation of the subventricular zone in multiple sclerosis: evidence for early glial progenitors. Proc Natl Acad Sci USA 104, 4694–4699. Abstract Abstract
- Nakatomi, H. Kuriu, T. Okabe, S. Yamamoto, S. Hatano, O. Kawahara, N. Tamura, A. Kirino, T. Nakafuku, M. (2002). Regeneration of hippocampal pyramidal neurons after ischemic brain injury by recruitment of endogenous neural progenitors. Cell 110, 429–441. Abstract Abstract
- Newton, S. S. Collier, E. F. Hunsberger, J. Adams, D. Terwilliger, R. Selvanayagam, E. Duman, R. S. (2003). Gene profile of electroconvulsive seizures: induction of neurotrophic and angiogenic factors. J Neurosci 23, 10841–10851. Abstract Abstract
- Niewmierzycka, A. Mills, J. St-Arnaud, R. Dedhar, S. Reichardt, L. F. (2005). Integrin-linked kinase deletion from mouse cortex results in cortical lamination defects resembling cobblestone lissencephaly. J Neurosci 25, 7022–7031. Abstract Abstract
- Noles, S. R. Chenn, A. (2007). Cadherin inhibition of beta-catenin signalling regulates the proliferation and differentiation of neural precursor cells. Mol Cell Neurosci 35, 549–558. Abstract Abstract
- Novak, U. Kaye, A.H. (2000). Extracellular matrix and the brain: components and function. J Clin Neurosci 7, 280–290. Abstract Abstract
- Olsson, M. Bjorklund, A. Campbell, K. (1998). Early specification of striatal projection neurons and interneuronal subtypes in the lateral and medial ganglionic eminence. Neuroscience 84, 867–876. Abstract Abstract
- Overstreet Wadiche, L. Bromberg, D. A. Bensen, A. L. Westbrook, G. L. (2005). GABAergic signalling to newborn neurons in dentate gyrus. J Neurophysiol 94, 4528–4532. Abstract Abstract
- Packer, M. A. Stasiv, Y. Benraiss, A. Chmielnicki, E. Grinberg, A. Westphal, H. Goldman, S. A. Enikolopov, G. (2003). Nitric oxide negatively regulates mammalian adult neurogenesis. Proc Natl Acad Sci USA 100, 9566–9571. Abstract Abstract
- Palma, V. Ruiz i Altaba, A. (2004). Hedgehog-GLI signalling regulates the behavior of cells with stem cell properties in the developing neocortex. Development 131, 337–345. Abstract Abstract
- Palmer, T. D. Willhoite, A. R. Gage, F. H. (2000). Vascular niche for adult hippocampal neurogenesis. J Comp Neurol 425, 479–494. Abstract Abstract
- Pang, T. Atefy, R. Sheen, V. (2008). Malformations of cortical development. Neurologist 14, 181–191. Abstract Abstract
- Peretto, P. Dati, C. De Marchis, S. Kim, H. H. Ukhanova, M. Fasolo, A. Margolis, F. L. (2004). Expression of the secreted factors noggin and bone morphogenetic proteins in the subependymal layer and olfactory bulb of the adult mouse brain. Neuroscience 128, 685–696. Abstract Abstract
- Phillips, W. Morton, A. J. Barker, R. A. (2005). Abnormalities of neurogenesis in the R6/2 mouse model of Huntington's disease are attributable to the in vivo microenvironment. J Neurosci 25, 11564–11576. Abstract Abstract
- Philpot, J. Pennock, J. Cowan, F. Sewry, C. A. Dubowitz, V. Bydder, G. Muntoni, F. (2000). Brain magnetic resonance imaging abnormalities in merosin-positive congenital muscular dystrophy. Eur J Paediatr Neurol 4, 109–114. Abstract Abstract
- Picard-Riera, N. Decker, L. Delarasse, C. Goude, K. Nait-Oumesmar, B. Liblau, R. Pham-Dinh, D. Evercooren, A. B. (2002). Experimental autoimmune encephalomyelitis mobilizes neural progenitors from the subventricular zone to undergo oligodendrogenesis in adult mice. Proc Natl Acad Sci USA 99, 13211–13216. Abstract Abstract
- Pierani, A. Brenner-Morton, S. Chiang, C. Jessell, T. M. (1999). A sonic hedgehog-independent, retinoid-activated pathway of neurogenesis in the ventral spinal cord. Cell 97, 903–915. Abstract Abstract
- Pilz, D. T. Matsumoto, N. Minnerath, S. Mills, P. Gleeson, J. G. Allen, K. M. Walsh, C. A. Barkovich, A. J. Dobyns, W. B. Ledbetter, D. H. (1998). LIS1 and XLIS (DCX) mutations cause most classical lissencephaly, but different patterns of malformation. Hum Mol Genet 7, 2029–2037. Abstract Abstract
- Pinnock, S. B. Balendra, R. Chan, M. Hunt, L. T. Turner-Stokes, T. Herbert, J. (2007). Interactions between nitric oxide and corticosterone in the regulation of progenitor cell proliferation in the dentate gyrus of the adult rat. Neuropsychopharmacology 32, 493–504. Abstract Abstract
- Pinto, L. Gotz, M. (2007). Radial glial cell heterogeneity–the source of diverse progeny in the CNS. Prog Neurobiol 83, 2–23. Abstract Abstract
- Plachta, N. Bibel, M. Tucker, K. L. Barde, Y. A. (2004). Developmental potential of defined neural progenitors derived from mouse embryonic stem cells. Development 131, 5449–5456. Abstract Abstract
- Qian, X. Goderie, S. K. Shen, Q. Stern, J. H. Temple, S. (1998). Intrinsic programs of patterned cell lineages in isolated vertebrate CNS ventricular zone cells. Development 125, 3143–3152. Abstract Abstract
- Qian, X. Shen, Q. Goderie, S. K. He, W. Capela, A. Davis, A. A. Temple, S. (2000). Timing of CNS cell generation: a programmed sequence of neuron and glial cell production from isolated murine cortical stem cells. Neuron 28, 69–80. Abstract Abstract
- Radice, G. L. Rayburn, H. Matsunami, H. Knudsen, K. A. Takeichi, M. Hynes, R. O. (1997). Developmental defects in mouse embryos lacking N-cadherin. Dev Biol 181, 64–78. Abstract Abstract
- Radley, J. J. Jacobs, B. L. (2002). 5-HT1A receptor antagonist administration decreases cell proliferation in the dentate gyrus. Brain Res 955, 264–267. Abstract Abstract
- Rakic, P. (2003). Developmental and evolutionary adaptations of cortical radial glia. Cereb Cortex 13, 541–549. Abstract Abstract
- Ramirez-Castillejo, C. Sanchez-Sanchez, F. Andreu-Agullo, C. Ferron, S. R. Aroca-Aguilar, J. D. Sanchez, P. Mira, H. Escribano, J. Farinas, I. (2006). Pigment epithelium-derived factor is a niche signal for neural stem cell renewal. Nat Neurosci 9, 331–339. Abstract Abstract
- Rauch, U. (1997). Modeling an extracellular environment for axonal pathfinding and fasciculation in the central nervous system. Cell Tissue Res 290, 349–356. Abstract Abstract
- Reiner, O. Carrozzo, R. Shen, Y. Wehnert, M. Faustinella, F. Dobyns, W. B. Caskey, C. T. Ledbetter, D. H. (1993). Isolation of a Miller-Dieker lissencephaly gene containing G protein beta-subunit-like repeats. Nature 364, 717–721. Abstract Abstract
- Riquelme, P. A. Drapeau, E. Doetsch, F. (2008). Brain micro-ecologies: neural stem cell niches in the adult mammalian brain. Philos Trans R Soc Lond B Biol Sci 363, 123–137. Abstract Abstract
- Rohatgi, R. Milenkovic, L. Scott, M. P. (2007). Patched1 regulates hedgehog signalling at the primary cilium. Science 317, 372–376. Abstract Abstract
- Ruoslahti, E. (1996). Brain extracellular matrix. Glycobiology 6, 489–492. Abstract Abstract
- Santarelli, L. Saxe, M. Gross, C. Surget, A. Battaglia, F. Dulawa, S. Weisstaub, N. Lee, J. Duman, R. Arancio, O. (2003). Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 301, 805–809. Abstract Abstract
- Sawamoto, K. Wichterle, H. Gonzalez-Perez, O. Cholfin, J. A. Yamada, M. Spassky, N. Murcia, N. S. Garcia-Verdugo, J. M. Marin, O. Rubenstein, J. L. (2006). New neurons follow the flow of cerebrospinal fluid in the adult brain. Science 311, 629–632. Abstract Abstract
- Scharfman, H. Goodman, J. Macleod, A. Phani, S. Antonelli, C. Croll, S. (2005). Increased neurogenesis and the ectopic granule cells after intrahippocampal BDNF infusion in adult rats. Exp Neurol 192, 348–356. Abstract Abstract
- Seaberg, R. M. van der Kooy, D. (2002). Adult rodent neurogenic regions: the ventricular subependyma contains neural stem cells, but the dentate gyrus contains restricted progenitors. J Neurosci 22, 1784–1793. Abstract Abstract
- Seri, B. Garcia-Verdugo, J. M. McEwen, B. S. Alvarez-Buylla, A. (2001). Astrocytes give rise to new neurons in the adult mammalian hippocampus. J Neurosci 21, 7153–7160. Abstract Abstract
- Seri, B. Garcia-Verdugo, J. M. Collado-Morente, L. McEwen, B. S. Alvarez-Buylla, A. (2004). Cell types, lineage, and architecture of the germinal zone in the adult dentate gyrus. J Comp Neurol 478, 359–378. Abstract Abstract
- Shen, Q. Goderie, S. K. Jin, L. Karanth, N. Sun, Y. Abramova, N. Vincent, P. Pumiglia, K. Temple, S. (2004). Endothelial cells stimulate self-renewal and expand neurogenesis of neural stem cells. Science 304, 1338–1340. Abstract Abstract
- Shen, Q. Wang, Y. Dimos, J. T. Fasano, C. A. Phoenix, T. N. Lemischka, I. R. Ivanova, N. B. Stifani, S. Morrisey, E. E. Temple, S. (2006). The timing of cortical neurogenesis is encoded within lineages of individual progenitor cells. Nat Neurosci 9, 743–751. Abstract Abstract
- Shimogori, T. Banuchi, V. Ng, H. Y. Strauss, J. B. Grove, E. A. (2004). Embryonic signalling centers expressing BMP, WNT and FGF proteins interact to pattern the cerebral cortex. Development 131, 5639–5647. Abstract Abstract
- Shors, T. J. Miesegaes, G. Beylin, A. Zhao, M. Rydel, T. Gould, E. (2001). Neurogenesis in the adult is involved in the formation of trace memories. Nature 410, 372–376. Abstract Abstract
- Singla, V. Reiter, J. F. (2006). The primary cilium as the cell's antenna: signalling at a sensory organelle. Science 313, 629–633. Abstract Abstract
- Sirko, S. von Holst, A. Wizenmann, A. Gotz, M. Faissner, A. (2007). Chondroitin sulfate glycosaminoglycans control proliferation, radial glia cell differentiation and neurogenesis in neural stem/progenitor cells. Development 134, 2727–2738. Abstract Abstract
- Smart, I. H. (1972). Proliferative characteristics of the ependymal layer during the early development of the spinal cord in the mouse. J Anat 111, 365–380. Abstract Abstract
- Storm, E. E. Rubenstein, J. L. Martin, G. R. (2003). Dosage of Fgf8 determines whether cell survival is positively or negatively regulated in the developing forebrain. Proc Natl Acad Sci USA 100, 1757–1762. Abstract Abstract
- Stuckmann, I. Weigmann, A. Shevchenko, A. Mann, M. Huttner, W. B. (2001). Ephrin B1 is expressed on neuroepithelial cells in correlation with neocortical neurogenesis. J Neurosci 21, 2726–2737. Abstract Abstract
- Stump, G. Durrer, A. Klein, A. L. Lutolf, S. Suter, U. Taylor, V. (2002). Notch1 and its ligands Delta-like and Jagged are expressed and active in distinct cell populations in the postnatal mouse brain. Mech Dev 114, 153–159. Abstract Abstract
- Takahashi, H. Liu, F. C. (2006). Genetic patterning of the mammalian telencephalon by morphogenetic molecules and transcription factors. Birth Defects Res C Embryo Today 78, 256–266. Abstract Abstract
- Thomas, L. B. Gates, M. A. Steindler, D. A. (1996). Young neurons from the adult subependymal zone proliferate and migrate along an astrocyte, extracellular matrix-rich pathway. Glia 17, 1–14. Abstract Abstract
- Thored, P. Wood, J. Arvidsson, A. Cammenga, J. Kokaia, Z. Lindvall, O. (2007). Long-term neuroblast migration along blood vessels in an area with transient angiogenesis and increased vascularization after stroke. Stroke 38, 3032–3039. Abstract Abstract
- Toda, T. Ikegawa, S. Okui, K. Kondo, E. Saito, K. Fukuyama, Y. Yoshioka, M. Kumagai, T. Suzumori, K. Kanazawa, I. (1994). Refined mapping of a gene responsible for Fukuyama-type congenital muscular dystrophy: evidence for strong linkage disequilibrium. Am J Hum Genet 55, 946–950. Abstract Abstract
- Vaccarino, F. M. Schwartz, M. L. Raballo, R. Nilsen, J. Rhee, J. Zhou, M. Doetschman, T. Coffin, J. D. Wyland, J. J. Hung, Y. T. (1999). Changes in cerebral cortex size are governed by fibroblast growth factor during embryogenesis. Nat Neurosci 2, 246–253. Abstract Abstract
- Van Raay, T. J. Wang, Y. K. Stark, M. R. Rasmussen, J. T. Francke, U. Vetter, M. L. Rao, M. S. (2001). frizzled 9 is expressed in neural precursor cells in the developing neural tube. Dev Genes Evol 211, 453–457. Abstract Abstract
- Vasudevan, A. Long, J. E. Crandall, J. E. Rubenstein, J. L. Bhide, P. G. (2008). Compartment-specific transcription factors orchestrate angiogenesis gradients in the embryonic brain. Nat Neurosci 11, 429–439. Abstract Abstract
- Vescovi, A. L. Reynolds, B. A. Fraser, D. D. Weiss, S. (1993). bFGF regulates the proliferative fate of unipotent (neuronal) and bipotent (neuronal/astroglial) EGF-generated CNS progenitor cells. Neuron 11, 951–966. Abstract Abstract
- Wada, K. Nakajima, A. Katayama, K. Kudo, C. Shibuya, A. Kubota, N. Terauchi, Y. Tachibana, M. Miyoshi, H. Kamisaki, Y. (2006). Peroxisome proliferator-activated receptor gamma-mediated regulation of neural stem cell proliferation and differentiation. J Biol Chem 281, 12673–12681. Abstract Abstract
- Wagner, J. P. Black, I. B. DiCicco-Bloom, E. (1999). Stimulation of neonatal and adult brain neurogenesis by subcutaneous injection of basic fibroblast growth factor. J Neurosci 19, 6006–6016. Abstract Abstract
- Walker, M. R. Stappenbeck, T. S. (2008). Deciphering the ‘black box’ of the intestinal stem cell niche: taking direction from other systems. Curr Opin Gastroenterol 24, 115–120. Abstract Abstract
- Wang, D. D. Krueger, D. D. Bordey, A. (2003). GABA depolarizes neuronal progenitors of the postnatal subventricular zone via GABAA receptor activation. J Physiol 550, 785–800. Abstract Abstract
- Wang, L. P. Kempermann, G. Kettenmann, H. (2005). A subpopulation of precursor cells in the mouse dentate gyrus receives synaptic GABAergic input. Mol Cell Neurosci 29, 181–189. Abstract Abstract
- Wechsler-Reya, R. J. Scott, M. P. (1999). Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog. Neuron 22, 103–114. Abstract Abstract
- Williams, B. P. Price, J. (1995). Evidence for multiple precursor cell types in the embryonic rat cerebral cortex. Neuron 14, 1181–1188. Abstract Abstract
- Woodhead, G. J. Mutch, C. A. Olson, E. C. Chenn, A. (2006). Cell-autonomous beta-catenin signalling regulates cortical precursor proliferation. J Neurosci 26, 12620–12630. Abstract Abstract
- Woods, C. G. Bond, J. Enard, W. (2005). Autosomal recessive primary microcephaly (MCPH): a review of clinical, molecular, and evolutionary findings. Am J Hum Genet 76, 717–728. Abstract Abstract
- Wrobel, C. N. Mutch, C. A. Swaminathan, S. Taketo, M. M. Chenn, A. (2007). Persistent expression of stabilized beta-catenin delays maturation of radial glial cells into intermediate progenitors. Dev Biol 309, 285–297. Abstract Abstract
- Yin, T. Li, L. (2006). The stem cell niches in bone. J Clin Invest 116, 1195–1201. Abstract Abstract
- Yoon, K. Gaiano, N. (2005). Notch signalling in the mammalian central nervous system: insights from mouse mutants. Nat Neurosci 8, 709–715. Abstract Abstract
- Yoshida, A. Kobayashi, K. Manya, H. Taniguchi, K. Kano, H. Mizuno, M. Inazu, T. Mitsuhashi, H. Takahashi, S. Takeuchi, M. (2001). Muscular dystrophy and neuronal migration disorder caused by mutations in a glycosyltransferase, POMGnT1. Dev Cell 1, 717–724. Abstract Abstract
- Yun, K. Fischman, S. Johnson, J. Hrabe de Angelis, M. Weinmaster, G. Rubenstein, J. L. (2002). Modulation of the notch signalling by Mash1 and Dlx1/2 regulates sequential specification and differentiation of progenitor cell types in the subcortical telencephalon. Development 129, 5029–5040. Abstract Abstract
- Zahs, K. R. (1998). Heterotypic coupling between glial cells of the mammalian central nervous system. Glia 24, 85–96. Abstract Abstract
- Zawadzka, M. Franklin, R. J. (2007). Myelin regeneration in demyelinating disorders: new developments in biology and clinical pathology. Curr Opin Neurol 20, 294–298. Abstract Abstract
- Zhang, R. Zhang, Z. Wang, L. Wang, Y. Gousev, A. Zhang, L. Ho, K. L. Morshead, C. Chopp, M. (2004). Activated neural stem cells contribute to stroke-induced neurogenesis and neuroblast migration toward the infarct boundary in adult rats. J Cereb Blood Flow Metab 24, 441–448. Abstract Abstract
- Zhang, R. L. Zhang, Z. G. Roberts, C. LeTourneau, Y. Lu, M. Zhang, L. Wang, Y. Chopp, M. (2008). Lengthening the G(1) phase of neural progenitor cells is concurrent with an increase of symmetric neuron generating division after stroke. J Cereb Blood Flow Metab 28, 602–611. Abstract Abstract
- de Chevigny, A. Lemasson, M. Saghatelyan, A. Sibbe, M. Schachner, M. Lledo, P. M. (2006). Delayed onset of odor detection in neonatal mice lacking tenascin-C. Mol Cell Neurosci 32, 174–186. Abstract Abstract
- von Holst, A. Sirko, S. Faissner, A. (2006). The unique 473HD-Chondroitinsulfate epitope is expressed by radial glia and involved in neural precursor cell proliferation. J Neurosci 26, 4082–4094. Abstract Abstract